 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
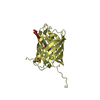
Yorodumi- PDB-1ggr: COMPLEX OF ENZYME IIAGLC AND THE HISTIDINE-CONTAINING PHOSPHOCARR... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 1ggr | ||||||
|---|---|---|---|---|---|---|---|
| Title | COMPLEX OF ENZYME IIAGLC AND THE HISTIDINE-CONTAINING PHOSPHOCARRIER PROTEIN HPR FROM ESCHERICHIA COLI NMR, RESTRAINED REGULARIZED MEAN STRUCTURE | ||||||
 Components Components |
| ||||||
 Keywords Keywords |  TRANSFERASE / PHOSPHOTRANSFERASE / KINASE / SUGAR TRANSPORT / COMPLEX (TRANSFERASE-PHOSPHOCARRIER) TRANSFERASE / PHOSPHOTRANSFERASE / KINASE / SUGAR TRANSPORT / COMPLEX (TRANSFERASE-PHOSPHOCARRIER) | ||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of carbohydrate metabolic process / phosphotransferase activity, nitrogenous group as acceptor / regulation of carbon utilization / antisigma factor binding / negative regulation of maltose transport / enzyme IIA-maltose transporter complex / negative regulation of transmembrane transport / positive regulation of glycogen catabolic process / phosphoenolpyruvate-dependent sugar phosphotransferase system / transmembrane transporter complex ...negative regulation of carbohydrate metabolic process / phosphotransferase activity, nitrogenous group as acceptor / regulation of carbon utilization / antisigma factor binding / negative regulation of maltose transport / enzyme IIA-maltose transporter complex / negative regulation of transmembrane transport / positive regulation of glycogen catabolic process / phosphoenolpyruvate-dependent sugar phosphotransferase system / transmembrane transporter complex / enzyme inhibitor activity / enzyme activator activity / enzyme regulator activity / kinase activity / phosphorylation / membrane / metal ion binding / cytosolSimilarity search - Function | ||||||
| Biological species |  Escherichia coli (E. coli) Escherichia coli (E. coli) | ||||||
| Method | SOLUTION NMR / RIGID BODY MINIMIZATION, CONSTRAINED, RESTRAINED SIMULATED ANNEALING | ||||||
 Authors Authors | Clore, G.M. / Wang, G. | ||||||
 Citation Citation | Journal: EMBO J. / Year: 2000 Title: Solution structure of the phosphoryl transfer complex between the signal transducing proteins HPr and IIA(glucose) of the Escherichia coli phosphoenolpyruvate:sugar phosphotransferase system. Authors: Wang, G. / Louis, J.M. / Sondej, M. / Seok, Y.J. / Peterkofsky, A. / Clore, G.M. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 1ggr.cif.gz | 227.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb1ggr.ent.gz | 191.7 KB | Display | PDB format |
| PDBx/mmJSON format | 1ggr.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/gg/1ggrftp://data.pdbj.org/pub/pdb/validation_reports/gg/1ggr | HTTPS FTP |
|---|
-Related structure data
| Related structure data | |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj- Assembly
Assembly
| Deposited unit | 
| |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| |||||||||
| NMR ensembles |
|
-Components
| #1: Protein | Mass: 18141.834 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Escherichia coli (E. coli) / Strain: GI698 / Production host: Escherichia coli (E. coli)References: UniProt: P69783, protein-Npi-phosphohistidine-sugar phosphotransferase |
|---|---|
| #2: Protein | Mass: 9129.332 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Escherichia coli (E. coli) / Strain: GI698 / Production host: Escherichia coli (E. coli) / References: UniProt: P0AA04 |
| #3: Chemical | ChemComp-PO3 / Phosphite ester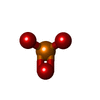  Mass: 78.972 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: PO3 Mass: 78.972 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: PO3 |
-Experimental details
-Experiment
| Experiment | Method: SOLUTION NMR |
|---|---|
| NMR details | Text: THE FOLLOWING EXPERIMENTS WERE CONDUCTED: (1) TRIPLE RESONANCE FOR ASSIGNMENT OF PROTEIN; (2) QUANTITATIVE J CORRELATION FOR COUPLING CONSTANTS; (3) 3D AND 4D HETERONUCLEAR SEPARATED AND ...Text: THE FOLLOWING EXPERIMENTS WERE CONDUCTED: (1) TRIPLE RESONANCE FOR ASSIGNMENT OF PROTEIN; (2) QUANTITATIVE J CORRELATION FOR COUPLING CONSTANTS; (3) 3D AND 4D HETERONUCLEAR SEPARATED AND FILTERED NOE EXPERIMENTS; (4) IPAP EXPERIMENTS FOR DIPOLAR COUPLINGS. DIPOLAR COUPLINGS WERE MEASURED IN A NEMATIC PHASE OF A COLLOIDAL SUSPENSION OF TMV (CLORE ET AL. 1998 J.AM.CHEM.SOC. 120, 105-106). |
- Sample preparation
Sample preparation
| Sample conditions | Ionic strength: 10 mM SODIUM PHOSPHATE / pH: 7.1 / Temperature: 308 K |
|---|---|
| Crystal grow | *PLUS Method: other / Details: NMR |
-NMR measurement
| NMR spectrometer |
|
|---|
- Processing
Processing
| NMR software | Name: X-PLOR / Version: NIH / Developer: Brunger / Classification: refinement |
|---|---|
| Refinement | Method: RIGID BODY MINIMIZATION, CONSTRAINED, RESTRAINED SIMULATED ANNEALING Software ordinal: 1 Details: THE STRUCTURES WERE CALCULATED BY RIGID BODY MINIMIZATION (CLORE (2000) PROC.NATL.ACAD. SCI. 97, 9021-9025; BEWLEY AND CLORE (2000) J.AM.CHEM.SOC. 122, 6009-6016) FOLLOWED BY ...Details: THE STRUCTURES WERE CALCULATED BY RIGID BODY MINIMIZATION (CLORE (2000) PROC.NATL.ACAD. SCI. 97, 9021-9025; BEWLEY AND CLORE (2000) J.AM.CHEM.SOC. 122, 6009-6016) FOLLOWED BY CONSTRAINED/RESTRAINED SIMULATED ANNEALING TO REFINE THE INTERFACIAL SIDECHAIN POSITIONS AND FINE TUNE THE RELATIVE ORIENTATION OF THE TWO PROTEINS (WANG ET AL. (2000) EMBO J. IN PRESS). THE TARGET FUNCTIONS COMPRISES TERMS FOR THE NOE RESTRAINTS, THE DIPOLAR COUPLING RESTRAINTS (CLORE ET AL. J.MAGN.RESON. 131, 159-162 (1998); J.MAGN.RESON. 133, 216-221(1998)), THE RADIUS OF GYRATION (KUSZEWSKI ET AL. (1999), AND A QUARTIC VAN DER WAALS REPULSION TERM (NILGES ET AL. (1988) FEBS LETT. 229, 129-136). THE STARTING COORDINATES COME FROM THE X-RAY STRUCTURES (WITH PROTONS ADDED) OF E. COLI HPR (1POH, JIA ET AL. (1993) J.BIOL.CHEM. 268, 22940-22501; RESOLUTION 1.5 A) AND IIAGLC (MOLECULE 2 OF 2F3G, FEESE ET AL. BIOCHEMISTRY 36, 16087-16096; RESOLUTION 2.0 A) IN SEVERAL DIFFERENT ORIENTATIONS WITH THE CA-CA DISTANCE BETWEEN THE ACTIVE SITE HISTIDINES RANGING FROM 28 TO 95 A, INCLUDING ORIENTATIONS WHERE THE TWO ACTIVE SITE HISTIDINES ARE NOT OPPOSED AND WHERE HPR IS DIRECTED TOWARDS THE FACE OF IIAGLC OPPOSITE TO THE IIAGLC ACTIVE SITE. ONLY THE INTERFACIAL SIDECHAINS ARE ALLOWED TO ALTER THEIR CONFORMATION; THE BACKBONE AND NON-INTERFACIAL SIDECHAINS OF ONE MOLECULE (IIAGLC) ARE HELD COMPLETELY FIXED; THE SECOND MOLECULE (HPR) CAN ROTATE AND TRANSLATE BUT THE RELATIVE COORDINATES OF ITS BACKBONE AND NON-INTERFACIAL SIDECHAINS ARE HELD FIXED. IN THIS ENTRY THE LAST COLUMN REPRESENTS THE AVERAGE RMS DIFFERENCE BETWEEN THE INDIVIDUAL SIMULATED ANNEALING STRUCTURES AND THE MEAN COORDINATE POSITIONS. IT IS IMPORTANT TO NOTE THAT THE VALUES GIVEN FOR THE BACKBONE ATOMS AND NON-INTERFACIAL SIDECHAINS PROVIDE ONLY A MEASURE OF THE PRECISION WITH WHICH THE RELATIVE OF THE TWO PROTEINS HAVE BEEN DETERMINED AND DOES NOT TAKE INTO ACCOUNT THE ERRORS IN THE X-RAY COORDINATES OF HPR AND IIAGLC. THREE SETS OF COORDINATES ARE GIVEN: MODEL 1: RESTRAINED MINIMIZED MEAN COORDINATES OF THE UNPHOSPHORYLATED HPR-IIAGLC COMPLEX SOLVED ON THE BASIS OF 82 INTERMOLECULAR DISTANCE RESTRAINTS (74 NOE DERIVED INTERPROTON DISTANCE AND 8 AMBIGUOUS INTERMOLECULAR SALT BRIDGE RESTRAINTS), 12 INTRAMOLECULAR INTERPROTON DISTANCE RESTRAINTS (RELATED SPECIFICALLY TO NOES INVOLVING RESIDUES 315 AND 317 OF HPR), 61 NMR DERIVED SIDECHAIN TORSION ANGLE RESTRAINTS, AND 195 1DNH DIPOLAR COUPLINGS (118 FOR IIAGLC AND 77 FOR HPR). CROSS-VALIDATION WAS USED FOR THE DIPOLAR COUPLINGS (CLORE AND GARRETT (1999) J. AM. CHEM. SOC. 121, 9008-9012). MODEL 2: RESTRAINED MINIMIZED MEAN COORDINATES FOR THE MODEL OF THE DISSOCIATIVE PHOSPHORYL TRANSITION STATE HPR-IIAGLC COMPLEX. EXPERIMENTAL RESTRAINTS ARE TO THOSE USED FOR MODEL 1, EXCEPT THAT ONE INTRAMOLECULAR INTERPROTON DISTANCE RESTRAINT INVOLVING HIS15 WAS REMOVED TO PERMIT A TRANSITION STATE TO FORM. IN ADDITION, COVALENT GEOMETRY RESTRAINTS ARE INCLUDED RELATING TO THE TRIGONAL BIPYRAMIDAL AT THE PHOSPHORUS. NO DISTANCE RESTRAINT IS INCLUDED FOR THE N-P BOND LENGTHS. THE CA-CA DISTANCE BETWEEN HIS315 (HPR) and HIS90 (IIAGLC) REMAINS UNCHANGED FROM MODEL 1, BUT THE ND1-NE2 DISTANCE BETWEEN HIS315 AND HIS90 IS REDUCED TO 6 A, WITH ESSENTIALLY IDEALIZED GEOMETRY OF THE PHOSPHORYL TRANSITION STATE. THE ND1-NE2 DISTANCE CORRESPONDS TO A DISSOCIATIVE TRANSITION STATE. THE RMS DIFFERENCE BETWEEN THE MEAN STRUCTURES OF THE UNPHOSPHORYLATED COMPLEX (MODEL 1) AND THE TRANSITION STATE COMPLEX IS 0.03 A FOR THE BACKBONE ATOMS AND 0.2 A FOR THE INTERFACIAL SIDECHAINS (EXCLUDING HIS315 AND HIS90). MODEL 3: RESTRAINED MINIMIZED MEAN COORDINATES FOR THE MODEL OF THE ASSOCIATIVE PHOSPHORYL TRANSITION STATE HPR-IIAGLC COMPLEX. MODEL 3 IS DERIVED FROM MODEL 2 BY CONSTRAINED/RESTRAINED MINIMIZATION IN WHICH THE COORDINATES OF ALL BACKBONE ATOMS, WITH THE OF RESIDUES 313-317 OF HPR AND RESIDUES 89-91 OF IIAGLC, AND ALL NON-INTERFACIAL SIDECHAINS ARE HELD COMPLETELY FIXED, AND IN WHICH THE N-P DISTANCES ARE RESTRAINED TO CA. 2 A, CORRESPONDING TO AN SN2 ASSOCIATIVE TRANSITION STATE. HPR-IIAGLC COMPLEX DEVIATIONS FROM IDEALIZED GEOMETRY: BONDS 0.014 A, ANGLES 1.74 A, IMPROPER TORSIONS 1.66 A RMS DEVIATIONS FROM NOE DISTANCE RESTRAINTS: 0.057 A RMS DEVIATIONS FROM SIDECHAIN TORSION ANGLE RESTRAINTS: 0.16 DEG. DIPOLAR COUPLING R-FACTORS (CLORE AND GARRETT (1999) J. AM. CHEM. SOC. 121, 9008-9012): 16.9% FOR HPR and 15.2% FOR IIAGLC (NOTE ONLY ONE ALIGNMENT TENSOR IS USED FOR BOTH HPR AND IIAGLC; FOR REFERENCE THE DIPOLAR COUPLING R-FACTORS FOR THE FREE X-RAY STRUCTURES OF HPR AND IIAGLC (USING INDIVIDUAL ALIGNMENT TENSORS FOR THE TWO PROTEINS) ARE 16.7% and 15.0%, RESPECTIVELY). |
| NMR ensemble | Conformer selection criteria: REGULARIZED MEAN STRUCTURES / Conformers calculated total number: 30 / Conformers submitted total number: 3 |
