 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
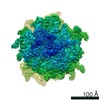
| 登録情報 | データベース: PDB / ID: 6sgc | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Rabbit 80S ribosome stalled on a poly(A) tail | ||||||||||||||||||
 要素 要素 |
| ||||||||||||||||||
 キーワード キーワード |  RIBOSOME (リボソーム) / Protein Translation (翻訳 (生物学)) / Ribosome Stalling / polyA tail RIBOSOME (リボソーム) / Protein Translation (翻訳 (生物学)) / Ribosome Stalling / polyA tail | ||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / DNA-(apurinic or apyrimidinic site) lyase / cytosolic ribosome / 5S rRNA binding / cytosolic large ribosomal subunit / リボソーム / structural constituent of ribosome / 翻訳 (生物学) / ribonucleoprotein complex / 核小体 ...maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / DNA-(apurinic or apyrimidinic site) lyase / cytosolic ribosome / 5S rRNA binding / cytosolic large ribosomal subunit / リボソーム / structural constituent of ribosome / 翻訳 (生物学) / ribonucleoprotein complex / 核小体 / RNA binding / 細胞質類似検索 - 分子機能 | ||||||||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) Oryctolagus cuniculus (ウサギ) Oryctolagus cuniculus (ウサギ) | ||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.8 Å | ||||||||||||||||||
 データ登録者 データ登録者 | Chandrasekaran, V. / Juszkiewicz, S. / Choi, J. / Puglisi, J.D. / Brown, A. / Shao, S. / Ramakrishnan, V. / Hegde, R.S. | ||||||||||||||||||
| 資金援助 |  英国, 英国,  米国, 5件 米国, 5件
| ||||||||||||||||||
 引用 引用 | ジャーナル: Nat Struct Mol Biol / 年: 2019 タイトル: Mechanism of ribosome stalling during translation of a poly(A) tail. 著者: Viswanathan Chandrasekaran / Szymon Juszkiewicz / Junhong Choi / Joseph D Puglisi / Alan Brown / Sichen Shao / V Ramakrishnan / Ramanujan S Hegde / 要旨: Faulty or damaged messenger RNAs are detected by the cell when translating ribosomes stall during elongation and trigger pathways of mRNA decay, nascent protein degradation and ribosome recycling. ...Faulty or damaged messenger RNAs are detected by the cell when translating ribosomes stall during elongation and trigger pathways of mRNA decay, nascent protein degradation and ribosome recycling. The most common mRNA defect in eukaryotes is probably inappropriate polyadenylation at near-cognate sites within the coding region. How ribosomes stall selectively when they encounter poly(A) is unclear. Here, we use biochemical and structural approaches in mammalian systems to show that poly-lysine, encoded by poly(A), favors a peptidyl-transfer RNA conformation suboptimal for peptide bond formation. This conformation partially slows elongation, permitting poly(A) mRNA in the ribosome's decoding center to adopt a ribosomal RNA-stabilized single-stranded helix. The reconfigured decoding center clashes with incoming aminoacyl-tRNA, thereby precluding elongation. Thus, coincidence detection of poly-lysine in the exit tunnel and poly(A) in the decoding center allows ribosomes to detect aberrant mRNAs selectively, stall elongation and trigger downstream quality control pathways essential for cellular homeostasis. | ||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6sgc.cif.gz | 4.8 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6sgc.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 6sgc.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/sg/6sgcftp://data.pdbj.org/pub/pdb/validation_reports/sg/6sgc | HTTPS FTP |
|---|
-関連構造データ


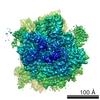
-リンク
 PDBj
PDBj






























- 集合体
集合体
| 登録構造単位 | 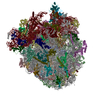
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 6種, 7分子 A1i15474842333
| #1: RNA鎖 | 分子量: 602777.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) |
|---|---|
| #35: RNA鎖 | 分子量: 3247.100 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主:  Salmonella virus SP6 (ウイルス) Salmonella virus SP6 (ウイルス) |
| #80: RNA鎖 | 分子量: 1167622.625 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) |
| #81: RNA鎖 | 5SリボソームRNA 分子量: 38691.914 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) |
| #82: RNA鎖 | 5.8SリボソームRNA 分子量: 50143.648 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) |
| #85: RNA鎖 | 分子量: 24776.012 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) |
+タンパク質 , 49種, 49分子 B1D1E1L1O1P1Q1S1T1U1V1W1Y1Z1a1b1e1h1B2C2F2G2H2L2O2P2Q2R2S2T2...
-40S ribosomal protein ... , 8種, 8分子 C1F1H1I1J1N1c1f1
| #3: タンパク質 | 分子量: 30002.061 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SS70 |
|---|---|
| #6: タンパク質 | 分子量: 29654.869 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TK17 |
| #8: タンパク質 | 分子量: 28751.906 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TM55 |
| #9: タンパク質 | 分子量: 22168.914 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SVB0 |
| #10: タンパク質 | 分子量: 24263.387 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TJW1 |
| #14: タンパク質 | 分子量: 14538.987 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SFR8 |
| #29: タンパク質 | 分子量: 9480.186 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TZ76 |
| #32: タンパク質 | 分子量: 14498.884 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1T8A2 |
-Ribosomal protein ... , 15種, 15分子 G1K1M1R1X1d1g1A2J2M2N2V2Y2j2t2
| #7: タンパク質 | 分子量: 22913.453 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TFM5 |
|---|---|
| #11: タンパク質 | リボソーム 分子量: 22641.564 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: B7NZS8 |
| #13: タンパク質 | 分子量: 18468.826 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TRM4 |
| #18: タンパク質 | 分子量: 16477.377 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SGX4 |
| #24: タンパク質 | リボソーム 分子量: 14865.555 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TG89 |
| #30: タンパク質 | 分子量: 7855.052 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TIB4 |
| #33: タンパク質 | リボソーム 分子量: 18004.041 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SK22 |
| #36: タンパク質 | 分子量: 28088.863 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TT27 |
| #45: タンパク質 | 分子量: 20288.465 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TUB8 |
| #47: タンパク質 | 分子量: 23870.549 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SZ12 |
| #48: タンパク質 | 分子量: 24207.285 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1T0C1 |
| #56: タンパク質 | 分子量: 14892.505 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1T6D1 |
| #59: タンパク質 | 分子量: 17303.363 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SQH0 |
| #70: タンパク質 | 分子量: 11111.032 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: U3KPD5 |
| #79: タンパク質 | 分子量: 17847.619 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SMR7 |
-60S ribosomal protein ... , 6種, 6分子 D2E2I2Z2i2n2
| #39: タンパク質 | 分子量: 34481.828 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SYJ6 |
|---|---|
| #40: タンパク質 | 分子量: 33028.336 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1SKF7 |
| #44: タンパク質 | / Ribosomal protein L10 (Predicted) 分子量: 24643.057 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: B7NZQ2 |
| #60: タンパク質 | 分子量: 15835.831 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TXF6 |
| #69: タンパク質 | 分子量: 12263.834 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: G1TTQ5 |
| #74: タンパク質・ペプチド | / eL41 分子量: 3473.451 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) / 参照: UniProt: A0A087WNH4 |
-タンパク質・ペプチド , 1種, 1分子 XX
| #83: タンパク質・ペプチド | 分子量: 2084.892 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Oryctolagus cuniculus (ウサギ) |
|---|
-非ポリマー , 3種, 285分子 

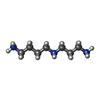
| #86: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 275 / 由来タイプ: 天然 / 式: Mg 分子量: 24.305 Da / 分子数: 275 / 由来タイプ: 天然 / 式: Mg#87: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn#88: 化合物 | スペルミジン 分子量: 145.246 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C7H19N3 分子量: 145.246 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C7H19N3 |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 4.5 MDa / 実験値: NO | ||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||
| 由来(組換発現) | 生物種: Salmonella virus SP6 (ウイルス) | ||||||||||||||||||||||||
| 緩衝液 | pH: 7.4 | ||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS | |||||||||||||||
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM | |||||||||||||||
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy | |||||||||||||||
| 撮影 |
|
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.8 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 148615 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL | ||||||||||||||||||||||||||||||||
| 精密化 | 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||||
| 拘束条件 |
|
