 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6emk | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
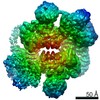
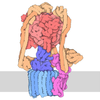
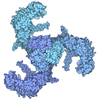
| タイトル | Cryo-EM Structure of Saccharomyces cerevisiae Target of Rapamycin Complex 2 | |||||||||||||||
 要素 要素 |
| |||||||||||||||
 キーワード キーワード |  SIGNALING PROTEIN / target of rapamycin (MTOR) / torc2 / FRB domain / Tor2-Lst8 / kinases SIGNALING PROTEIN / target of rapamycin (MTOR) / torc2 / FRB domain / Tor2-Lst8 / kinases | |||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報PIP3 activates AKT signaling / CD28 dependent PI3K/Akt signaling / regulation of snRNA pseudouridine synthesis / mitochondria-nucleus signaling pathway / Regulation of TP53 Degradation / positive regulation of Rho guanyl-nucleotide exchange factor activity / TORC2 signaling / 1-phosphatidylinositol 4-kinase / 1-phosphatidylinositol 4-kinase activity / establishment or maintenance of actin cytoskeleton polarity ...PIP3 activates AKT signaling / CD28 dependent PI3K/Akt signaling / regulation of snRNA pseudouridine synthesis / mitochondria-nucleus signaling pathway / Regulation of TP53 Degradation / positive regulation of Rho guanyl-nucleotide exchange factor activity / TORC2 signaling / 1-phosphatidylinositol 4-kinase / 1-phosphatidylinositol 4-kinase activity / establishment or maintenance of actin cytoskeleton polarity / VEGFR2 mediated vascular permeability / HSF1-dependent transactivation / fungal-type cell wall organization / TORC2 complex / Amino acids regulate mTORC1 / TORC1 complex / fungal-type vacuole membrane / vacuolar membrane / positive regulation of Rho protein signal transduction / negative regulation of macroautophagy / positive regulation of endocytosis / MTOR / cytoskeleton organization / phosphatidylinositol-4,5-bisphosphate binding / nuclear periphery / negative regulation of autophagy / response to nutrient / regulation of cell growth / regulation of actin cytoskeleton organization / リボソーム生合成 / endosome membrane / regulation of cell cycle / non-specific serine/threonine protein kinase / 細胞周期 / ゴルジ体 / protein phosphorylation / protein serine kinase activity / protein serine/threonine kinase activity / protein-containing complex binding / シグナル伝達 / ミトコンドリア / ATP binding / 細胞核 / 細胞膜 / 細胞質基質 / 細胞質類似検索 - 分子機能 | |||||||||||||||
| 生物種 |  Saccharomyces cerevisiae (パン酵母) Saccharomyces cerevisiae (パン酵母) | |||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 7.9 Å | |||||||||||||||
 データ登録者 データ登録者 | Karuppasamy, M. / Kusmider, B. / Oliveira, T.M. / Gaubitz, C. / Prouteau, M. / Loewith, R. / Schaffitzel, C. | |||||||||||||||
| 資金援助 | 4件
| |||||||||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2017 タイトル: Cryo-EM structure of Saccharomyces cerevisiae target of rapamycin complex 2. 著者: Manikandan Karuppasamy / Beata Kusmider / Taiana M Oliveira / Christl Gaubitz / Manoel Prouteau / Robbie Loewith / Christiane Schaffitzel /    要旨: The target of rapamycin (TOR) kinase assembles into two distinct multiprotein complexes, conserved across eukaryote evolution. In contrast to TOR complex 1 (TORC1), TORC2 kinase activity is not ...The target of rapamycin (TOR) kinase assembles into two distinct multiprotein complexes, conserved across eukaryote evolution. In contrast to TOR complex 1 (TORC1), TORC2 kinase activity is not inhibited by the macrolide rapamycin. Here, we present the structure of Saccharomyces cerevisiae TORC2 determined by electron cryo-microscopy. TORC2 contains six subunits assembling into a 1.4 MDa rhombohedron. Tor2 and Lst8 form the common core of both TOR complexes. Avo3/Rictor is unique to TORC2, but interacts with the same HEAT repeats of Tor2 that are engaged by Kog1/Raptor in mammalian TORC1, explaining the mutual exclusivity of these two proteins. Density, which we conclude is Avo3, occludes the FKBP12-rapamycin-binding site of Tor2's FRB domain rendering TORC2 rapamycin insensitive and recessing the kinase active site. Although mobile, Avo1/hSin1 further restricts access to the active site as its conserved-region-in-the-middle (CRIM) domain is positioned along an edge of the TORC2 active-site-cleft, consistent with a role for CRIM in substrate recruitment. #1: ジャーナル: Mol Cell / 年: 2015タイトル: Molecular Basis of the Rapamycin Insensitivity of Target Of Rapamycin Complex 2. 著者: Christl Gaubitz / Taiana M Oliveira / Manoel Prouteau / Alexander Leitner / Manikandan Karuppasamy / Georgia Konstantinidou / Delphine Rispal / Sandra Eltschinger / Graham C Robinson / ...著者: Christl Gaubitz / Taiana M Oliveira / Manoel Prouteau / Alexander Leitner / Manikandan Karuppasamy / Georgia Konstantinidou / Delphine Rispal / Sandra Eltschinger / Graham C Robinson / Stéphane Thore / Ruedi Aebersold / Christiane Schaffitzel / Robbie Loewith / 要旨: Target of Rapamycin (TOR) plays central roles in the regulation of eukaryote growth as the hub of two essential multiprotein complexes: TORC1, which is rapamycin-sensitive, and the lesser ...Target of Rapamycin (TOR) plays central roles in the regulation of eukaryote growth as the hub of two essential multiprotein complexes: TORC1, which is rapamycin-sensitive, and the lesser characterized TORC2, which is not. TORC2 is a key regulator of lipid biosynthesis and Akt-mediated survival signaling. In spite of its importance, its structure and the molecular basis of its rapamycin insensitivity are unknown. Using crosslinking-mass spectrometry and electron microscopy, we determined the architecture of TORC2. TORC2 displays a rhomboid shape with pseudo-2-fold symmetry and a prominent central cavity. Our data indicate that the C-terminal part of Avo3, a subunit unique to TORC2, is close to the FKBP12-rapamycin-binding domain of Tor2. Removal of this sequence generated a FKBP12-rapamycin-sensitive TORC2 variant, which provides a powerful tool for deciphering TORC2 function in vivo. Using this variant, we demonstrate a role for TORC2 in G2/M cell-cycle progression. | |||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6emk.cif.gz | 1.2 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6emk.ent.gz | 1001 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6emk.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/em/6emkftp://data.pdbj.org/pub/pdb/validation_reports/em/6emk | HTTPS FTP |
|---|
-関連構造データ





-リンク
 PDBj
PDBj




- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 281915.438 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (strain ATCC 204508 / S288c) (パン酵母)遺伝子: TOR2, DRR2, TSC14, YKL203C / 発現宿主: Saccharomyces cerevisiae (パン酵母)参照: UniProt: P32600, 1-phosphatidylinositol 4-kinase, non-specific serine/threonine protein kinase#2: タンパク質 | MTOR / TORC subunit LST8 / Lethal with SEC13 protein 8分子量: 34077.879 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (strain ATCC 204508 / S288c) (パン酵母)遺伝子: LST8, YNL006W, N2005 / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: P41318#3: タンパク質 | 分子量: 25804.662 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (strain ATCC 204508 / S288c) (パン酵母)発現宿主: Saccharomyces cerevisiae (パン酵母)#4: タンパク質 | 分子量: 47206.457 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (strain ATCC 204508 / S288c) (パン酵母)遺伝子: AVO2, YMR068W, YM9916.07 / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: Q04749#5: タンパク質 | 分子量: 131565.453 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (strain ATCC 204508 / S288c) (パン酵母)遺伝子: AVO1, YOL078W, O1110 / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: Q08236 |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Target of rapamycin protein complex 2MTOR / タイプ: COMPLEX / Entity ID: all / 由来: RECOMBINANT |
|---|---|
| 分子量 | 値: 1.4 MDa / 実験値: NO |
| 由来(天然) | 生物種: Saccharomyces cerevisiae (strain ATCC 204508 / S288c) (パン酵母) |
| 由来(組換発現) | 生物種: Saccharomyces cerevisiae (パン酵母) |
| 緩衝液 | pH: 7.5 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R2/2 |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 4 K / 詳細: 2 - 3 sec blotting |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS | |||||||||||||||||||||
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM | |||||||||||||||||||||
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy / 倍率(公称値): 105000 X / 最大 デフォーカス(公称値): 3500 nm / 最小 デフォーカス(公称値): 1500 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: COMA FREE | |||||||||||||||||||||
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER 最高温度: 70 K / 最低温度: 70 K | |||||||||||||||||||||
| 撮影 | Imaging-ID: 1
| |||||||||||||||||||||
| 画像スキャン | 横: 3710 / 縦: 3838 / 動画フレーム数/画像: 40 / 利用したフレーム数/画像: 1-40 |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0158 / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 画像処理 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | PDB-ID: 5FVM Pdb chain residue range: 81-2474 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 8→220.8 Å / Cor.coef. Fo:Fc: 0.987 / SU B: 111.293 / SU ML: 0.794 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 溶媒モデル: PARAMETERS FOR MASK CACLULATION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 295.421 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 合計: 48016 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|
