 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5o9g | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Structure of nucleosome-Chd1 complex | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード |  DNA BINDING PROTEIN (DNA結合タンパク質) / ATPase (ATPアーゼ) / Complex / Nucleosome (ヌクレオソーム) / DNA (デオキシリボ核酸) / Chromatin Remodeling (クロマチンリモデリング) DNA BINDING PROTEIN (DNA結合タンパク質) / ATPase (ATPアーゼ) / Complex / Nucleosome (ヌクレオソーム) / DNA (デオキシリボ核酸) / Chromatin Remodeling (クロマチンリモデリング) | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報nucleolar chromatin / regulation of transcriptional start site selection at RNA polymerase II promoter / negative regulation of DNA-templated DNA replication / regulation of chromatin organization / nucleosome organization / rDNA binding / SLIK (SAGA-like) complex / SAGA complex / ATP-dependent chromatin remodeler activity / sister chromatid cohesion ...nucleolar chromatin / regulation of transcriptional start site selection at RNA polymerase II promoter / negative regulation of DNA-templated DNA replication / regulation of chromatin organization / nucleosome organization / rDNA binding / SLIK (SAGA-like) complex / SAGA complex / ATP-dependent chromatin remodeler activity / sister chromatid cohesion / termination of RNA polymerase II transcription / termination of RNA polymerase I transcription / ATP-dependent activity, acting on DNA / methylated histone binding / helicase activity / transcription elongation by RNA polymerase II / 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 / chromatin DNA binding / structural constituent of chromatin / ヌクレオソーム / histone binding / transcription cis-regulatory region binding / クロマチンリモデリング / protein heterodimerization activity / chromatin binding / クロマチン / regulation of transcription by RNA polymerase II / ATP hydrolysis activity / ミトコンドリア / DNA binding / 核質 / ATP binding / 細胞核類似検索 - 分子機能 | ||||||
| 生物種 | Xenopus laevis (アフリカツメガエル) Saccharomyces cerevisiae S288c (パン酵母) Saccharomyces cerevisiae S288c (パン酵母)synthetic construct (人工物) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.8 Å | ||||||
 データ登録者 データ登録者 | Farnung, L. / Vos, S.M. / Wigge, C. / Cramer, P. | ||||||
| 資金援助 |  ドイツ, 1件 ドイツ, 1件
| ||||||
 引用 引用 | ジャーナル: Nature / 年: 2017 タイトル: Nucleosome-Chd1 structure and implications for chromatin remodelling. 著者: Lucas Farnung / Seychelle M Vos / Christoph Wigge / Patrick Cramer / 要旨: Chromatin-remodelling factors change nucleosome positioning and facilitate DNA transcription, replication, and repair. The conserved remodelling factor chromodomain-helicase-DNA binding protein ...Chromatin-remodelling factors change nucleosome positioning and facilitate DNA transcription, replication, and repair. The conserved remodelling factor chromodomain-helicase-DNA binding protein 1(Chd1) can shift nucleosomes and induce regular nucleosome spacing. Chd1 is required for the passage of RNA polymerase IIthrough nucleosomes and for cellular pluripotency. Chd1 contains the DNA-binding domains SANT and SLIDE, a bilobal motor domain that hydrolyses ATP, and a regulatory double chromodomain. Here we report the cryo-electron microscopy structure of Chd1 from the yeast Saccharomyces cerevisiae bound to a nucleosome at a resolution of 4.8 Å. Chd1 detaches two turns of DNA from the histone octamer and binds between the two DNA gyres in a state poised for catalysis. The SANT and SLIDE domains contact detached DNA around superhelical location (SHL) -7 of the first DNA gyre. The ATPase motor binds the second DNA gyre at SHL +2 and is anchored to the N-terminal tail of histone H4, as seen in a recent nucleosome-Snf2 ATPase structure. Comparisons with published results reveal that the double chromodomain swings towards nucleosomal DNA at SHL +1, resulting in ATPase closure. The ATPase can then promote translocation of DNA towards the nucleosome dyad, thereby loosening the first DNA gyre and remodelling the nucleosome. Translocation may involve ratcheting of the two lobes of the ATPase, which is trapped in a pre- or post-translocation state in the absence or presence, respectively, of transition state-mimicking compounds. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5o9g.cif.gz | 490.9 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5o9g.ent.gz | 377.4 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5o9g.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/o9/5o9gftp://data.pdbj.org/pub/pdb/validation_reports/o9/5o9g | HTTPS FTP |
|---|
-関連構造データ


-リンク
 PDBj
PDBj






































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 4種, 7分子 AEBFCGW
| #1: タンパク質 | 分子量: 15435.126 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Xenopus laevis (アフリカツメガエル)発現宿主:  Escherichia coli (大腸菌) / 参照: UniProt: P84233 Escherichia coli (大腸菌) / 参照: UniProt: P84233#2: タンパク質 | ヒストンH4分子量: 11394.426 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Xenopus laevis (アフリカツメガエル)発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P62799#3: タンパク質 | ヒストンH2A分子量: 14109.436 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Xenopus laevis (アフリカツメガエル)遺伝子: hist1h2aj, LOC494591 / 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: Q6AZJ8, UniProt: P06897*PLUS#8: タンパク質 | | 分子量: 168496.609 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae S288c (パン酵母)遺伝子: CHD1, YER164W, SYGP-ORF4 / 発現宿主:  Trichoplusia ni (イラクサキンウワバ) Trichoplusia ni (イラクサキンウワバ)参照: UniProt: P32657, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 |
|---|
-Histone H2B 1. ... , 2種, 2分子 DH
| #4: タンパク質 | 分子量: 13655.948 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) Xenopus laevis (アフリカツメガエル)発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P02281 |
|---|---|
| #5: タンパク質 | 分子量: 13526.769 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) Xenopus laevis (アフリカツメガエル)発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P02281 |
-DNA鎖 , 2種, 2分子 IJ
| #6: DNA鎖 | 分子量: 64143.914 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|---|
| #7: DNA鎖 | 分子量: 64293.945 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
-非ポリマー , 2種, 2分子 
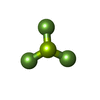
| #9: 化合物 | ChemComp-ADP / アデノシン二リン酸 分子量: 427.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM |
|---|---|
| #10: 化合物 | ChemComp-BEF /  分子量: 66.007 Da / 分子数: 1 / 由来タイプ: 合成 / 式: BeF3 分子量: 66.007 Da / 分子数: 1 / 由来タイプ: 合成 / 式: BeF3 |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER |
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy / 最大 デフォーカス(公称値): 2750 nm / 最小 デフォーカス(公称値): 1250 nm / Cs: 2.7 mm / C2レンズ絞り径: 150 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 10 sec. / 電子線照射量: 3 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 実像数: 3802 |
| 電子光学装置 | エネルギーフィルター名称: GIF Quantum |
| 画像スキャン | 動画フレーム数/画像: 40 / 利用したフレーム数/画像: 1-40 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 990000 | ||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||
| 3次元再構成 | 解像度: 4.8 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 67032 / アルゴリズム: FOURIER SPACE / 対称性のタイプ: POINT |
