 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 4nu1 | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Crystal structure of a transition state mimic of the GSK-3/Axin complex bound to phosphorylated N-terminal auto-inhibitory pS9 peptide | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | TRANSFERASE/PEPTIDE / Wnt / LRP6 /  GSK-3 (GSK-3) / Axin (AXIN1) / kinase (キナーゼ) / primed substrate / transition state (遷移状態) / phosphorylated N-terminal auto-inhibitory pS9 peptide / TRANSFERASE-PEPTIDE complex GSK-3 (GSK-3) / Axin (AXIN1) / kinase (キナーゼ) / primed substrate / transition state (遷移状態) / phosphorylated N-terminal auto-inhibitory pS9 peptide / TRANSFERASE-PEPTIDE complex | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報hepatic stellate cell activation / B-WICH complex positively regulates rRNA expression / negative regulation of synaptic assembly at neuromuscular junction / negative regulation of neuron maturation / Regulation of HSF1-mediated heat shock response / negative regulation of protein localization to centrosome / re-entry into mitotic cell cycle / Beta-catenin phosphorylation cascade / CRMPs in Sema3A signaling / membrane-bounded organelle ...hepatic stellate cell activation / B-WICH complex positively regulates rRNA expression / negative regulation of synaptic assembly at neuromuscular junction / negative regulation of neuron maturation / Regulation of HSF1-mediated heat shock response / negative regulation of protein localization to centrosome / re-entry into mitotic cell cycle / Beta-catenin phosphorylation cascade / CRMPs in Sema3A signaling / membrane-bounded organelle / Disassembly of the destruction complex and recruitment of AXIN to the membrane / positive regulation of synaptic assembly at neuromuscular junction / myotube differentiation / positive regulation of osteoclast proliferation / armadillo repeat domain binding / negative regulation of neuron migration / cell growth involved in cardiac muscle cell development / positive regulation of cardiac muscle cell differentiation / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / head development / negative regulation of dendrite morphogenesis / Degradation of beta-catenin by the destruction complex / 細胞生物学 / neuron projection organization / regulation of microtubule anchoring at centrosome / dorsal/ventral axis specification / negative regulation of cardiac muscle hypertrophy / positive regulation of stem cell differentiation / negative regulation of mesenchymal stem cell differentiation / beta-catenin destruction complex disassembly / protein localization to microtubule / axial mesoderm formation / negative regulation of dendrite development / superior temporal gyrus development / positive regulation of protein localization to cilium / negative regulation of smooth muscle cell apoptotic process / GLI3 is processed to GLI3R by the proteasome / negative regulation of dopaminergic neuron differentiation / maintenance of cell polarity / positive regulation of protein localization to centrosome / autosome genomic imprinting / : / positive regulation of mitochondrial outer membrane permeabilization involved in apoptotic signaling pathway / 減数分裂 / positive regulation of cilium assembly / bone remodeling / myoblast fusion / post-anal tail morphogenesis / negative regulation of protein acetylation / negative regulation of TOR signaling / APC truncation mutants have impaired AXIN binding / AXIN missense mutants destabilize the destruction complex / Truncations of AMER1 destabilize the destruction complex / protein serine/threonine kinase binding / epigenetic programming in the zygotic pronuclei / beta-catenin destruction complex / positive regulation of ubiquitin-dependent protein catabolic process / tau-protein kinase / regulation of microtubule-based process / regulation of modification of postsynaptic structure / regulation of protein export from nucleus / Beta-catenin phosphorylation cascade / Signaling by GSK3beta mutants / CTNNB1 S33 mutants aren't phosphorylated / CTNNB1 S37 mutants aren't phosphorylated / CTNNB1 S45 mutants aren't phosphorylated / CTNNB1 T41 mutants aren't phosphorylated / positive regulation of osteoclast differentiation / axon extension / I-SMAD binding / negative regulation of protein metabolic process / cellular response to interleukin-3 / cellular response to glucocorticoid stimulus / Wnt signalosome / regulation of axon extension / positive regulation of mitochondrial membrane potential / regulation of long-term synaptic potentiation / 紡錘体 / negative regulation of protein localization to nucleus / Disassembly of the destruction complex and recruitment of AXIN to the membrane / nucleocytoplasmic transport / regulation of neurotransmitter receptor localization to postsynaptic specialization membrane / positive regulation of cell-matrix adhesion / negative regulation of calcineurin-NFAT signaling cascade / regulation of osteoblast differentiation / negative regulation of fat cell differentiation / establishment or maintenance of cell polarity / positive regulation of DNA biosynthetic process / cellular response to hepatocyte growth factor stimulus / dynein complex binding / regulation of neuronal synaptic plasticity / regulation of dendrite morphogenesis / regulation of axonogenesis / establishment of cell polarity / tau-protein kinase activity / response to zinc ion / glycogen metabolic process / RUNX1 regulates transcription of genes involved in WNT signaling / RUNX1 regulates estrogen receptor mediated transcription / ER overload response類似検索 - 分子機能 | ||||||
| 生物種 |  Mus musculus (ハツカネズミ) Mus musculus (ハツカネズミ) Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | X線回折 / シンクロトロン / 分子置換 / 解像度: 2.5 Å | ||||||
 データ登録者 データ登録者 | Chu, M.L.-H. / Stamos, J.L. / Enos, M.D. / Shah, N. / Weis, W.I. | ||||||
 引用 引用 | ジャーナル: Elife / 年: 2014 タイトル: Structural basis of GSK-3 inhibition by N-terminal phosphorylation and by the Wnt receptor LRP6. 著者: Stamos, J.L. / Chu, M.L. / Enos, M.D. / Shah, N. / Weis, W.I. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 4nu1.cif.gz | 174.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb4nu1.ent.gz | 138.2 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 4nu1.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/nu/4nu1ftp://data.pdbj.org/pub/pdb/validation_reports/nu/4nu1 | HTTPS FTP |
|---|
-関連構造データ







-リンク
 PDBj
PDBj








- 集合体
集合体
| 登録構造単位 | 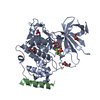
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| 単位格子 |
|
-要素
-タンパク質 / タンパク質・ペプチド , 2種, 2分子 AB
| #1: タンパク質 | / GSK-3 beta / Serine/threonine-protein kinase GSK3B 分子量: 44260.605 Da / 分子数: 1 / 断片: Residues 1-383 with phosphoylated Ser9 / 由来タイプ: 組換発現 / 由来: (組換発現) Mus musculus (ハツカネズミ) / 遺伝子: Gsk3b / プラスミド: pET29b(+) / 発現宿主:  Escherichia coli (大腸菌) / 株 (発現宿主): BL21(DE3)Codon-plus RIL Escherichia coli (大腸菌) / 株 (発現宿主): BL21(DE3)Codon-plus RIL参照: UniProt: Q9WV60, tau-protein kinase, non-specific serine/threonine protein kinase |
|---|---|
| #2: タンパク質・ペプチド | AXIN1 / Axis inhibition protein 1 / hAxin 分子量: 2738.144 Da / 分子数: 1 / 断片: Residues 383-402 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: AXIN1, AXIN / プラスミド: Modified pGEX-KG / 発現宿主: Escherichia coli (大腸菌) / 株 (発現宿主): BL21(DE3)Codon-plus RIL / 参照: UniProt: O15169 |
-非ポリマー , 6種, 105分子 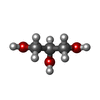


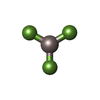
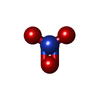

| #3: 化合物 | グリセリン 分子量: 92.094 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C3H8O3 分子量: 92.094 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C3H8O3#4: 化合物 |  分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg#5: 化合物 | ChemComp-ADP / | アデノシン二リン酸 分子量: 427.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM#6: 化合物 | ChemComp-AF3 / | フッ化アルミニウム 分子量: 83.977 Da / 分子数: 1 / 由来タイプ: 合成 / 式: AlF3 分子量: 83.977 Da / 分子数: 1 / 由来タイプ: 合成 / 式: AlF3#7: 化合物 | ChemComp-NO3 / | 硝酸塩 分子量: 62.005 Da / 分子数: 1 / 由来タイプ: 合成 / 式: NO3 分子量: 62.005 Da / 分子数: 1 / 由来タイプ: 合成 / 式: NO3#8: 水 | ChemComp-HOH / | 水分子量: 18.015 Da / 分子数: 98 / 由来タイプ: 天然 / 式: H2O |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.83 Å3/Da / 溶媒含有率: 56.48 % |
|---|---|
| 結晶化 | 温度: 277 K / 手法: microdialysis / pH: 7.5 詳細: 10% PEG 35,000, 20mM Tris pH7.5, 300mM NaCl, 5% glycerol, 10mM MgCl2, 200uM ATP, and 5mM DTT, MICRODIALYSIS, temperature 277K |
-データ収集
| 回折 | 平均測定温度: 78 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: APS  / ビームライン: 23-ID-B / 波長: 1.033 Å / ビームライン: 23-ID-B / 波長: 1.033 Å |
| 検出器 | タイプ: MARMOSAIC 300 mm CCD / 検出器: CCD / 日付: 2013年11月15日 / 詳細: mirrors |
| 放射 | モノクロメーター: Si(III) / プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 1.033 Å / 相対比: 1 |
| 反射 | 解像度: 2.5→46.76 Å / Num. all: 19925 / Num. obs: 19922 / % possible obs: 100 % / Observed criterion σ(F): 0 / 冗長度: 21 % / Biso Wilson estimate: 60.85 Å2 / Rsym value: 0.303 / Net I/σ(I): 13 |
| 反射 シェル | 解像度: 2.5→2.6 Å / 冗長度: 21.6 % / Mean I/σ(I) obs: 0.7 / Rsym value: 6.772 / % possible all: 100 |
-位相決定
| 位相決定 | 手法: 分子置換 |
|---|
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: 4NM3 解像度: 2.5→40.498 Å / Occupancy max: 1 / Occupancy min: 0.38 / SU ML: 0.39 / σ(F): 1.35 / 位相誤差: 26.15 / 立体化学のターゲット値: ML
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 74.7733 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 2.5→40.498 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: X-RAY DIFFRACTION
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|
