ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
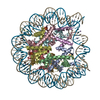
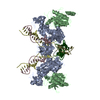
| 登録情報 | データベース: PDB / ID: 6rny | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
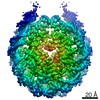
| タイトル | PFV intasome - nucleosome strand transfer complex | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード |  DNA BINDING PROTEIN (DNA結合タンパク質) / chromatin (クロマチン) / nucleosome (ヌクレオソーム) / retrovirus (レトロウイルス科) DNA BINDING PROTEIN (DNA結合タンパク質) / chromatin (クロマチン) / nucleosome (ヌクレオソーム) / retrovirus (レトロウイルス科) | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報nucleosomal DNA binding / 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素; アスパラギン酸プロテアーゼ / リボヌクレアーゼH / RNA polymerase II core promoter sequence-specific DNA binding / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine ...nucleosomal DNA binding / 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素; アスパラギン酸プロテアーゼ / リボヌクレアーゼH / RNA polymerase II core promoter sequence-specific DNA binding / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / Inhibition of DNA recombination at telomere / Meiotic synapsis / telomere organization / RNA Polymerase I Promoter Opening / SUMOylation of chromatin organization proteins / Assembly of the ORC complex at the origin of replication / DNAメチル化 / Condensation of Prophase Chromosomes / HCMV Late Events / Chromatin modifications during the maternal to zygotic transition (MZT) / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / SIRT1 negatively regulates rRNA expression / innate immune response in mucosa / PRC2 methylates histones and DNA / virion component / Defective pyroptosis / HDACs deacetylate histones / RNA Polymerase I Promoter Escape / Nonhomologous End-Joining (NHEJ) / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / NoRC negatively regulates rRNA expression / G2/M DNA damage checkpoint / B-WICH complex positively regulates rRNA expression / HDMs demethylate histones / DNA Damage/Telomere Stress Induced Senescence / Metalloprotease DUBs / PKMTs methylate histone lysines / RMTs methylate histone arginines / 遺伝的組換え / Pre-NOTCH Transcription and Translation / nucleosome assembly / Activation of anterior HOX genes in hindbrain development during early embryogenesis / HCMV Early Events / DNA integration / Transcriptional regulation of granulopoiesis / structural constituent of chromatin / 逆転写酵素 / viral genome integration into host DNA / viral penetration into host nucleus / establishment of integrated proviral latency / UCH proteinases / RNA-directed DNA polymerase activity / ヌクレオソーム / 転移酵素; リンを含む基を移すもの; 核酸を移すもの / antimicrobial humoral immune response mediated by antimicrobial peptide / RNA-DNA hybrid ribonuclease activity / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / RUNX1 regulates transcription of genes involved in differentiation of HSCs / chromatin organization / Factors involved in megakaryocyte development and platelet production / Processing of DNA double-strand break ends / HATs acetylate histones / antibacterial humoral response / Senescence-Associated Secretory Phenotype (SASP) / positive regulation of cell growth / Oxidative Stress Induced Senescence / DNA recombination / Estrogen-dependent gene expression / host cell cytoplasm / chromosome, telomeric region / 加水分解酵素; エステル加水分解酵素 / DNAポリメラーゼ / aspartic-type endopeptidase activity / DNA-directed DNA polymerase activity / Ub-specific processing proteases / defense response to Gram-positive bacterium / symbiont entry into host cell / RNA polymerase II cis-regulatory region sequence-specific DNA binding / Amyloid fiber formation / protein heterodimerization activity / host cell nucleus / enzyme binding / protein-containing complex / タンパク質分解 / DNA binding / extracellular space / RNA binding / extracellular exosome / extracellular region / 核質 / 生体膜 / identical protein binding類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) Human spumaretrovirus (ウイルス) Human spumaretrovirus (ウイルス) Pyrobaculum filamentous virus 1 (ウイルス) Pyrobaculum filamentous virus 1 (ウイルス) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.9 Å | |||||||||
 データ登録者 データ登録者 | Pye, V.E. / Renault, L. / Maskell, D.P. / Cherepanov, P. / Costa, A. | |||||||||
| 資金援助 |  英国, 2件 英国, 2件
| |||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2019 タイトル: Retroviral integration into nucleosomes through DNA looping and sliding along the histone octamer. 著者: Marcus D Wilson / Ludovic Renault / Daniel P Maskell / Mohamed Ghoneim / Valerie E Pye / Andrea Nans / David S Rueda / Peter Cherepanov / Alessandro Costa /  要旨: Retroviral integrase can efficiently utilise nucleosomes for insertion of the reverse-transcribed viral DNA. In face of the structural constraints imposed by the nucleosomal structure, integrase ...Retroviral integrase can efficiently utilise nucleosomes for insertion of the reverse-transcribed viral DNA. In face of the structural constraints imposed by the nucleosomal structure, integrase gains access to the scissile phosphodiester bonds by lifting DNA off the histone octamer at the site of integration. To clarify the mechanism of DNA looping by integrase, we determined a 3.9 Å resolution structure of the prototype foamy virus intasome engaged with a nucleosome core particle. The structural data along with complementary single-molecule Förster resonance energy transfer measurements reveal twisting and sliding of the nucleosomal DNA arm proximal to the integration site. Sliding the nucleosomal DNA by approximately two base pairs along the histone octamer accommodates the necessary DNA lifting from the histone H2A-H2B subunits to allow engagement with the intasome. Thus, retroviral integration into nucleosomes involves the looping-and-sliding mechanism for nucleosomal DNA repositioning, bearing unexpected similarities to chromatin remodelers. #1: ジャーナル: Nat Commun / 年: 2019タイトル: Retroviral integration into nucleosomes through DNA looping and sliding along the histone octamer. 著者: Marcus D Wilson / Ludovic Renault / Daniel P Maskell / Mohamed Ghoneim / Valerie E Pye / Andrea Nans / David S Rueda / Peter Cherepanov / Alessandro Costa / 要旨: Retroviral integrase can efficiently utilise nucleosomes for insertion of the reverse-transcribed viral DNA. In face of the structural constraints imposed by the nucleosomal structure, integrase ...Retroviral integrase can efficiently utilise nucleosomes for insertion of the reverse-transcribed viral DNA. In face of the structural constraints imposed by the nucleosomal structure, integrase gains access to the scissile phosphodiester bonds by lifting DNA off the histone octamer at the site of integration. To clarify the mechanism of DNA looping by integrase, we determined a 3.9 Å resolution structure of the prototype foamy virus intasome engaged with a nucleosome core particle. The structural data along with complementary single-molecule Förster resonance energy transfer measurements reveal twisting and sliding of the nucleosomal DNA arm proximal to the integration site. Sliding the nucleosomal DNA by approximately two base pairs along the histone octamer accommodates the necessary DNA lifting from the histone H2A-H2B subunits to allow engagement with the intasome. Thus, retroviral integration into nucleosomes involves the looping-and-sliding mechanism for nucleosomal DNA repositioning, bearing unexpected similarities to chromatin remodelers. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6rny.cif.gz | 478.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6rny.ent.gz | 369.3 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6rny.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/rn/6rnyftp://data.pdbj.org/pub/pdb/validation_reports/rn/6rny | HTTPS FTP |
|---|
-関連構造データ



-リンク
 PDBj
PDBj










































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 5種, 12分子 AEBFCGDHKLOP
| #1: タンパク質 | H3F3A 分子量: 15360.983 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: H3F3A, H3.3A, H3F3, PP781, H3F3B, H3.3B / 発現宿主:  Escherichia coli (大腸菌) / 参照: UniProt: P84243 Escherichia coli (大腸菌) / 参照: UniProt: P84243#2: タンパク質 | ヒストンH4分子量: 11394.426 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, ...遺伝子: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, H4FE, HIST1H4K, H4/D, H4FD, HIST1H4L, H4/K, H4FK, HIST2H4A, H4/N, H4F2, H4FN, HIST2H4, HIST2H4B, H4/O, H4FO, HIST4H4 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P62805#3: タンパク質 | 分子量: 14121.537 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H2AG, H2AFP, HIST1H2AI, H2AFC, HIST1H2AK, H2AFD, HIST1H2AL, H2AFI, HIST1H2AM, H2AFN 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P0C0S8#4: タンパク質 | 分子量: 13937.213 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H2BC, H2BFL, HIST1H2BE, H2BFH, HIST1H2BF, H2BFG, HIST1H2BG, H2BFA, HIST1H2BI, H2BFK 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P62807#5: タンパク質 | インテグラーゼ / Pr125Pol分子量: 44456.695 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Human spumaretrovirus (ウイルス) / 遺伝子: pol / 発現宿主: Escherichia coli (大腸菌)参照: UniProt: P14350, 逆転写酵素, DNAポリメラーゼ, リボヌクレアーゼH, 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素; ...参照: UniProt: P14350, 逆転写酵素, DNAポリメラーゼ, リボヌクレアーゼH, 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素; アスパラギン酸プロテアーゼ, 転移酵素; リンを含む基を移すもの; 核酸を移すもの, 加水分解酵素; エステル加水分解酵素 |
|---|
-DNA鎖 , 5種, 6分子 IUTJQM
| #6: DNA鎖 | 分子量: 39189.094 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
|---|---|
| #7: DNA鎖 | 分子量: 33581.453 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
| #8: DNA鎖 | 分子量: 10114.523 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
| #9: DNA鎖 | 分子量: 16397.531 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
| #10: DNA鎖 | 分子量: 5834.794 Da / 分子数: 2 / 由来タイプ: 合成 由来: (合成) Pyrobaculum filamentous virus 1 (ウイルス) |
-非ポリマー , 1種, 2分子 
| #11: 化合物 |  分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.399 MDa / 実験値: NO | ||||||||||||||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7 | ||||||||||||||||||||||||||||||||||||||||||
| 緩衝液成分 | 濃度: 1 MM / 名称: DTT | ||||||||||||||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.1 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 400 divisions/in. / グリッドのタイプ: C-flat-1/1 | ||||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: LEICA EM CPC / 凍結剤: ETHANE / 湿度: 80 % / 凍結前の試料温度: 277 K / 詳細: 1 min incubation 3.5s blot |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy / 倍率(公称値): 59000 X / 最大 デフォーカス(公称値): 3.5 nm / 最小 デフォーカス(公称値): 1.5 nm / Cs: 2.7 mm / C2レンズ絞り径: 70 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 1.6 sec. / 電子線照射量: 56 e/Å2 / 検出モード: INTEGRATING フィルム・検出器のモデル: FEI FALCON II (4k x 4k) 撮影したグリッド数: 2 / 実像数: 4916 |
| 電子光学装置 | 球面収差補正装置: Microscope was modified with a Cs corrector (NeCEN netherlands) |
| 画像スキャン | 動画フレーム数/画像: 7 |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.14_3235: / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 3125 | ||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||
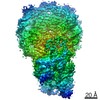
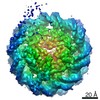
| 3次元再構成 | 解像度: 3.9 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 177155 / アルゴリズム: FOURIER SPACE / クラス平均像の数: 3 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | 空間: REAL 詳細: The initial model was placed in the density using Chimera. Manual building was performed in Coot and final refinement was carried out using phenix.real_space_refine and namdinator. Additional ...詳細: The initial model was placed in the density using Chimera. Manual building was performed in Coot and final refinement was carried out using phenix.real_space_refine and namdinator. Additional restraints describing protein secondary structure, DNA base pairing and stacking were used in Phenix. | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
|