 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8hnv | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | CryoEM structure of HpaCas9-sgRNA-dsDNA in the presence of AcrIIC4 | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | ANTIMICROBIAL PROTEIN / Cas9 / cleavage inhibition / HYDROLASE-RNA-ANTIMICROBIAL PROTEIN complex | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報maintenance of CRISPR repeat elements / endonuclease activity / defense response to virus / 加水分解酵素; エステル加水分解酵素 / DNA binding / RNA binding / metal ion binding 類似検索 - 分子機能 | ||||||
| 生物種 |  Haemophilus parainfluenzae (パラインフルエンザ菌) Haemophilus parainfluenzae (パラインフルエンザ菌)synthetic construct (人工物) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.1 Å | ||||||
 データ登録者 データ登録者 | Sun, W. / Cheng, Z. / Wang, J. / Yang, X. / Wang, Y. | ||||||
| 資金援助 |  中国, 1件 中国, 1件
| ||||||
 引用 引用 | ジャーナル: Proc Natl Acad Sci U S A / 年: 2023 タイトル: AcrIIC4 inhibits type II-C Cas9 by preventing R-loop formation. 著者: Wei Sun / Zhi Cheng / Jiuyu Wang / Jing Yang / Xueyan Li / Jinlong Wang / Minxuan Chen / Xiaoqi Yang / Gang Sheng / Jizhong Lou / Yanli Wang / 要旨: Anti-CRISPR (Acr) proteins are encoded by phages and other mobile genetic elements and inhibit host CRISPR-Cas immunity using versatile strategies. AcrIIC4 is a broad-spectrum Acr that inhibits the ...Anti-CRISPR (Acr) proteins are encoded by phages and other mobile genetic elements and inhibit host CRISPR-Cas immunity using versatile strategies. AcrIIC4 is a broad-spectrum Acr that inhibits the type II-C CRISPR-Cas9 system in several species by an unknown mechanism. Here, we determined a series of structures of Cas9 (HpaCas9)-sgRNA in complex with AcrIIC4 and/or target DNA, as well as the crystal structure of AcrIIC4 alone. We found that AcrIIC4 resides in the crevice between the REC1 and REC2 domains of HpaCas9, where its extensive interactions restrict the mobility of the REC2 domain and prevent the unwinding of target double-stranded (ds) DNA at the PAM-distal end. Therefore, the full-length guide RNA:target DNA heteroduplex fails to form in the presence of AcrIIC4, preventing Cas9 nuclease activation. Altogether, our structural and biochemical studies illuminate a unique Acr mechanism that allows DNA binding to the Cas9 effector complex but blocks its cleavage by preventing R-loop formation, a key step supporting DNA cleavage by Cas9. #1: ジャーナル: mBio / 年: 2018 タイトル: Potent Cas9 Inhibition in Bacterial and Human Cells by AcrIIC4 and AcrIIC5 Anti-CRISPR Proteins. 著者: Jooyoung Lee / Aamir Mir / Alireza Edraki / Bianca Garcia / Nadia Amrani / Hannah E Lou / Ildar Gainetdinov / April Pawluk / Raed Ibraheim / Xin D Gao / Pengpeng Liu / Alan R Davidson / Karen ...著者: Jooyoung Lee / Aamir Mir / Alireza Edraki / Bianca Garcia / Nadia Amrani / Hannah E Lou / Ildar Gainetdinov / April Pawluk / Raed Ibraheim / Xin D Gao / Pengpeng Liu / Alan R Davidson / Karen L Maxwell / Erik J Sontheimer /   要旨: In their natural settings, CRISPR-Cas systems play crucial roles in bacterial and archaeal adaptive immunity to protect against phages and other mobile genetic elements, and they are also widely used ...In their natural settings, CRISPR-Cas systems play crucial roles in bacterial and archaeal adaptive immunity to protect against phages and other mobile genetic elements, and they are also widely used as genome engineering technologies. Previously we discovered bacteriophage-encoded Cas9-specific anti-CRISPR (Acr) proteins that serve as countermeasures against host bacterial immunity by inactivating their CRISPR-Cas systems (A. Pawluk, N. Amrani, Y. Zhang, B. Garcia, et al., Cell 167:1829-1838.e9, 2016, https://doi.org/10.1016/j.cell.2016.11.017). We hypothesized that the evolutionary advantages conferred by anti-CRISPRs would drive the widespread occurrence of these proteins in nature (K. L. Maxwell, Mol Cell 68:8-14, 2017, https://doi.org/10.1016/j.molcel.2017.09.002; A. Pawluk, A. R. Davidson, and K. L. Maxwell, Nat Rev Microbiol 16:12-17, 2018, https://doi.org/10.1038/nrmicro.2017.120; E. J. Sontheimer and A. R. Davidson, Curr Opin Microbiol 37:120-127, 2017, https://doi.org/10.1016/j.mib.2017.06.003). We have identified new anti-CRISPRs using the same bioinformatic approach that successfully identified previous Acr proteins (A. Pawluk, N. Amrani, Y. Zhang, B. Garcia, et al., Cell 167:1829-1838.e9, 2016, https://doi.org/10.1016/j.cell.2016.11.017) against Cas9 (NmeCas9). In this work, we report two novel anti-CRISPR families in strains of and , both of which harbor type II-C CRISPR-Cas systems (A. Mir, A. Edraki, J. Lee, and E. J. Sontheimer, ACS Chem Biol 13:357-365, 2018, https://doi.org/10.1021/acschembio.7b00855). We characterize the type II-C Cas9 orthologs from and , show that the newly identified Acrs are able to inhibit these systems, and define important features of their inhibitory mechanisms. The Acr is the most potent NmeCas9 inhibitor identified to date. Although inhibition of NmeCas9 by anti-CRISPRs from and reveals cross-species inhibitory activity, more distantly related type II-C Cas9s are not inhibited by these proteins. The specificities of anti-CRISPRs and divergent Cas9s appear to reflect coevolution of their strategies to combat or evade each other. Finally, we validate these new anti-CRISPR proteins as potent off-switches for Cas9 genome engineering applications. As one of their countermeasures against CRISPR-Cas immunity, bacteriophages have evolved natural inhibitors known as anti-CRISPR (Acr) proteins. Despite the existence of such examples for type II CRISPR-Cas systems, we currently know relatively little about the breadth of Cas9 inhibitors, and most of their direct Cas9 targets are uncharacterized. In this work we identify two new type II-C anti-CRISPRs and their cognate Cas9 orthologs, validate their functionality and in bacteria, define their inhibitory spectrum against a panel of Cas9 orthologs, demonstrate that they act before Cas9 DNA binding, and document their utility as off-switches for Cas9-based tools in mammalian applications. The discovery of diverse anti-CRISPRs, the mechanistic analysis of their cognate Cas9s, and the definition of Acr inhibitory mechanisms afford deeper insight into the interplay between Cas9 orthologs and their inhibitors and provide greater scope for exploiting Acrs for CRISPR-based genome engineering. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8hnv.cif.gz | 235.8 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8hnv.ent.gz | 167.5 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8hnv.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/hn/8hnvftp://data.pdbj.org/pub/pdb/validation_reports/hn/8hnv | HTTPS FTP |
|---|
-関連構造データ

-リンク
 PDBj
PDBj































































- 集合体
集合体
| 登録構造単位 | 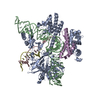
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 121605.695 Da / 分子数: 1 / 変異: D13A,H581A / 由来タイプ: 組換発現 詳細: Two mutations D13A and H581A were introduced to inactivate the catalytic sites of HpaCas9. The first residue 'Ser' of the sample sequence is the one expressed from the vector left after tag cleavage. 由来: (組換発現) Haemophilus parainfluenzae (パラインフルエンザ菌)遺伝子: csn1 / 発現宿主: |
|---|---|
| #2: RNA鎖 | 分子量: 40860.125 Da / 分子数: 1 / 由来タイプ: 合成 詳細: RNA is originally derived from Haemophilus parainfluenzae and modified by author. 由来: (合成) synthetic construct (人工物) |
| #3: DNA鎖 | 分子量: 10865.029 Da / 分子数: 1 / 由来タイプ: 合成 由来: (合成) Haemophilus parainfluenzae (パラインフルエンザ菌) |
| #4: DNA鎖 | 分子量: 10766.954 Da / 分子数: 1 / 由来タイプ: 合成 由来: (合成) Haemophilus parainfluenzae (パラインフルエンザ菌) |
| #5: タンパク質 | 分子量: 10072.395 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: WP_049372635.1;The first residue 'Ser' of the sample sequence is the one expressed from the vector left after tag cleavage. 由来: (組換発現) Haemophilus parainfluenzae (パラインフルエンザ菌)遺伝子: acrIIC4 / 発現宿主: |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.194 MDa / 実験値: YES | ||||||||||||||||||||||||
| 由来(天然) | 生物種: Haemophilus parainfluenzae (パラインフルエンザ菌) | ||||||||||||||||||||||||
| 由来(組換発現) | 生物種: | ||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 1600 nm / 最小 デフォーカス(公称値): 1000 nm |
| 撮影 | 電子線照射量: 60 e/Å2 フィルム・検出器のモデル: GATAN K2 QUANTUM (4k x 4k) |
- 解析
解析
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3次元再構成 | 解像度: 3.1 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 212049 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT | ||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 17.66 Å2 | ||||||||||||||||||||||||
| 拘束条件 |
|
