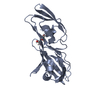
ジャーナル: PLoS Pathog / 年: 2023 タイトル: An unbroken network of interactions connecting flagellin domains is required for motility in viscous environments. 著者: Marko Nedeljković / Mark A B Kreutzberger / Sandra Postel / Daniel Bonsor / Yingying Xing / Neil Jacob / William J Schuler / Edward H Egelman / Eric J Sundberg / 要旨: In its simplest form, bacterial flagellar filaments are composed of flagellin proteins with just two helical inner domains, which together comprise the filament core. Although this minimal filament ...In its simplest form, bacterial flagellar filaments are composed of flagellin proteins with just two helical inner domains, which together comprise the filament core. Although this minimal filament is sufficient to provide motility in many flagellated bacteria, most bacteria produce flagella composed of flagellin proteins with one or more outer domains arranged in a variety of supramolecular architectures radiating from the inner core. Flagellin outer domains are known to be involved in adhesion, proteolysis and immune evasion but have not been thought to be required for motility. Here we show that in the Pseudomonas aeruginosa PAO1 strain, a bacterium that forms a ridged filament with a dimerization of its flagellin outer domains, motility is categorically dependent on these flagellin outer domains. Moreover, a comprehensive network of intermolecular interactions connecting the inner domains to the outer domains, the outer domains to one another, and the outer domains back to the inner domain filament core, is required for motility. This inter-domain connectivity confers PAO1 flagella with increased stability, essential for its motility in viscous environments. Additionally, we find that such ridged flagellar filaments are not unique to Pseudomonas but are, instead, present throughout diverse bacterial phyla.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード CELL ADHESION (細胞接着) /
CELL ADHESION (細胞接着) /  機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 引用
引用

 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード

 PDBj
PDBj 集合体
集合体

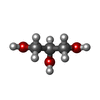
 分子量: 92.094 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C3H8O3
分子量: 92.094 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C3H8O3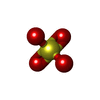
 分子量: 96.063 Da / 分子数: 2 / 由来タイプ: 合成 / 式: SO4
分子量: 96.063 Da / 分子数: 2 / 由来タイプ: 合成 / 式: SO4 分子量: 18.015 Da / 分子数: 346 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 346 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 解析
解析