 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-7mvw: Crystal structure of Chaetomium thermophilum Nup188 NTD (residues... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7mvw | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Crystal structure of Chaetomium thermophilum Nup188 NTD (residues 1-1134) | |||||||||||||||
 Components Components | Nucleoporin NUP188 | |||||||||||||||
 Keywords Keywords |  TRANSPORT PROTEIN / nuclear pore complex / nucleocytoplasmic transport / alpha-helical solenoid / nuclear pore TRANSPORT PROTEIN / nuclear pore complex / nucleocytoplasmic transport / alpha-helical solenoid / nuclear pore | |||||||||||||||
| Function / homology |  Function and homology information Function and homology informationstructural constituent of nuclear pore / mRNA transport / nuclear pore / protein transport / nuclear membraneSimilarity search - Function | |||||||||||||||
| Biological species |  Chaetomium thermophilum (fungus) Chaetomium thermophilum (fungus) | |||||||||||||||
| Method | X-RAY DIFFRACTION / SYNCHROTRON / MAD / Resolution: 2.76 Å | |||||||||||||||
 Authors Authors | Petrovic, S. / Samanta, D. / Perriches, T. / Bley, C.J. / Thierbach, K. / Brown, B. / Nie, S. / Mobbs, G.W. / Stevens, T.A. / Liu, X. ...Petrovic, S. / Samanta, D. / Perriches, T. / Bley, C.J. / Thierbach, K. / Brown, B. / Nie, S. / Mobbs, G.W. / Stevens, T.A. / Liu, X. / Tomaleri, G.P. / Schaus, L. / Hoelz, A. | |||||||||||||||
| Funding support |  United States, 4items United States, 4items
| |||||||||||||||
 Citation Citation | Journal: Science / Year: 2022 Title: Architecture of the linker-scaffold in the nuclear pore. Authors: Stefan Petrovic / Dipanjan Samanta / Thibaud Perriches / Christopher J Bley / Karsten Thierbach / Bonnie Brown / Si Nie / George W Mobbs / Taylor A Stevens / Xiaoyu Liu / Giovani Pinton ...Authors: Stefan Petrovic / Dipanjan Samanta / Thibaud Perriches / Christopher J Bley / Karsten Thierbach / Bonnie Brown / Si Nie / George W Mobbs / Taylor A Stevens / Xiaoyu Liu / Giovani Pinton Tomaleri / Lucas Schaus / André Hoelz / Abstract: INTRODUCTION In eukaryotic cells, the selective bidirectional transport of macromolecules between the nucleus and cytoplasm occurs through the nuclear pore complex (NPC). Embedded in nuclear envelope ...INTRODUCTION In eukaryotic cells, the selective bidirectional transport of macromolecules between the nucleus and cytoplasm occurs through the nuclear pore complex (NPC). Embedded in nuclear envelope pores, the ~110-MDa human NPC is an ~1200-Å-wide and ~750-Å-tall assembly of ~1000 proteins, collectively termed nucleoporins. Because of the NPC's eightfold rotational symmetry along the nucleocytoplasmic axis, each of the ~34 different nucleoporins occurs in multiples of eight. Architecturally, the NPC's symmetric core is composed of an inner ring encircling the central transport channel and two outer rings anchored on both sides of the nuclear envelope. Because of its central role in the flow of genetic information from DNA to RNA to protein, the NPC is commonly targeted in viral infections and its nucleoporin constituents are associated with a plethora of diseases. RATIONALE Although the arrangement of most scaffold nucleoporins in the NPC's symmetric core was determined by quantitative docking of crystal structures into cryo-electron tomographic (cryo-ET) maps of intact NPCs, the topology and molecular details of their cohesion by multivalent linker nucleoporins have remained elusive. Recently, in situ cryo-ET reconstructions of NPCs from various species have indicated that the NPC's inner ring is capable of reversible constriction and dilation in response to variations in nuclear envelope membrane tension, thereby modulating the diameter of the central transport channel by ~200 Å. We combined biochemical reconstitution, high-resolution crystal and single-particle cryo-electron microscopy (cryo-EM) structure determination, docking into cryo-ET maps, and physiological validation to elucidate the molecular architecture of the linker-scaffold interaction network that not only is essential for the NPC's integrity but also confers the plasticity and robustness necessary to allow and withstand such large-scale conformational changes. RESULTS By biochemically mapping scaffold-binding regions of all fungal and human linker nucleoporins and determining crystal and single-particle cryo-EM structures of linker-scaffold complexes, we completed the characterization of the biochemically tractable linker-scaffold network and established its evolutionary conservation, despite considerable sequence divergence. We determined a series of crystal and single-particle cryo-EM structures of the intact Nup188 and Nup192 scaffold hubs bound to their Nic96, Nup145N, and Nup53 linker nucleoporin binding regions, revealing that both proteins form distinct question mark-shaped keystones of two evolutionarily conserved hetero‑octameric inner ring complexes. Linkers bind to scaffold surface pockets through short defined motifs, with flanking regions commonly forming additional disperse interactions that reinforce the binding. Using a structure‑guided functional analysis in , we confirmed the robustness of linker‑scaffold interactions and established the physiological relevance of our biochemical and structural findings. The near-atomic composite structures resulting from quantitative docking of experimental structures into human and cryo-ET maps of constricted and dilated NPCs structurally disambiguated the positioning of the Nup188 and Nup192 hubs in the intact fungal and human NPC and revealed the topology of the linker-scaffold network. The linker-scaffold gives rise to eight relatively rigid inner ring spokes that are flexibly interconnected to allow for the formation of lateral channels. Unexpectedly, we uncovered that linker‑scaffold interactions play an opposing role in the outer rings by forming tight cross-link staples between the eight nuclear and cytoplasmic outer ring spokes, thereby limiting the dilatory movements to the inner ring. CONCLUSION We have substantially advanced the structural and biochemical characterization of the symmetric core of the and human NPCs and determined near-atomic composite structures. The composite structures uncover the molecular mechanism by which the evolutionarily conserved linker‑scaffold establishes the NPC's integrity while simultaneously allowing for the observed plasticity of the central transport channel. The composite structures are roadmaps for the mechanistic dissection of NPC assembly and disassembly, the etiology of NPC‑associated diseases, the role of NPC dilation in nucleocytoplasmic transport of soluble and integral membrane protein cargos, and the anchoring of asymmetric nucleoporins. [Figure: see text]. | |||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7mvw.cif.gz | 393.7 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7mvw.ent.gz | 320.5 KB | Display | PDB format |
| PDBx/mmJSON format | 7mvw.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/mv/7mvwftp://data.pdbj.org/pub/pdb/validation_reports/mv/7mvw | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  7mvtC  7mvuC  7mvvC  7mvxC  7mvyC  7mvzC  7mw0C  7mw1C  7tbiC  7tbjC  7tbkC C: citing same article ( |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj
- Assembly
Assembly
| Deposited unit | 
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 125400.016 Da / Num. of mol.: 1 / Fragment: N-terminal domain, UNP residues 1-1134 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Chaetomium thermophilum (strain DSM 1495 / CBS 144.50 / IMI 039719) (fungus)Strain: DSM 1495 / CBS 144.50 / IMI 039719 / Gene: NUP188, CTHT_0070850 / Production host:  Escherichia coli (E. coli) / References: UniProt: G0SFH5 Escherichia coli (E. coli) / References: UniProt: G0SFH5 | ||||
|---|---|---|---|---|---|
| #2: Chemical | Glycerol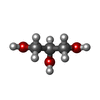  Mass: 92.094 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C3H8O3 Mass: 92.094 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C3H8O3#3: Water | ChemComp-HOH / | Water Mass: 18.015 Da / Num. of mol.: 34 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 34 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | N | |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 3.85 Å3/Da / Density % sol: 68.05 % |
|---|---|
| Crystal grow | Temperature: 294 K / Method: vapor diffusion, hanging drop / pH: 8 / Details: 10 % (w/v) PEG 3350, 0.1 M KSCN |
-Data collection
| Diffraction |
| ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diffraction source |
| ||||||||||||||||||
| Detector |
| ||||||||||||||||||
| Radiation |
| ||||||||||||||||||
| Radiation wavelength |
| ||||||||||||||||||
| Reflection | Resolution: 2.76→45 Å / Num. obs: 48981 / % possible obs: 99.8 % / Redundancy: 10.4 % / Biso Wilson estimate: 70 Å2 / CC1/2: 1 / CC star: 1 / Rmerge(I) obs: 0.162 / Rpim(I) all: 0.053 / Rrim(I) all: 0.171 / Net I/σ(I): 13.7 | ||||||||||||||||||
| Reflection shell | Resolution: 2.76→2.859 Å / Redundancy: 9.5 % / Rmerge(I) obs: 2.103 / Mean I/σ(I) obs: 1.26 / Num. unique obs: 4880 / CC1/2: 0.57 / CC star: 0.85 / Rpim(I) all: 0.715 / Rrim(I) all: 2.225 / % possible all: 99.63 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MAD / Resolution: 2.76→43.39 Å / SU ML: 0.4059 / Cross valid method: FREE R-VALUE / σ(F): 1.36 / Phase error: 30.2914 Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 81.38 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.76→43.39 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
