Ministry of Education, Youth and Sports of the Czech Republic
LM2018127
チェコ
引用
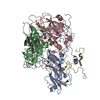
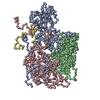
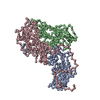
ジャーナル: Commun Biol / 年: 2022 タイトル: Cryo-electron microscopy and image classification reveal the existence and structure of the coxsackievirus A6 virion. 著者: Carina R Büttner / Radovan Spurný / Tibor Füzik / Pavel Plevka / 要旨: Coxsackievirus A6 (CV-A6) has recently overtaken enterovirus A71 and CV-A16 as the primary causative agent of hand, foot, and mouth disease worldwide. Virions of CV-A6 were not identified in previous ...Coxsackievirus A6 (CV-A6) has recently overtaken enterovirus A71 and CV-A16 as the primary causative agent of hand, foot, and mouth disease worldwide. Virions of CV-A6 were not identified in previous structural studies, and it was speculated that the virus is unique among enteroviruses in using altered particles with expanded capsids to infect cells. In contrast, the virions of other enteroviruses are required for infection. Here we used cryo-electron microscopy (cryo-EM) to determine the structures of the CV-A6 virion, altered particle, and empty capsid. We show that the CV-A6 virion has features characteristic of virions of other enteroviruses, including a compact capsid, VP4 attached to the inner capsid surface, and fatty acid-like molecules occupying the hydrophobic pockets in VP1 subunits. Furthermore, we found that in a purified sample of CV-A6, the ratio of infectious units to virions is 1 to 500. Therefore, it is likely that virions of CV-A6 initiate infection, like those of other enteroviruses. Our results provide evidence that future vaccines against CV-A6 should target its virions instead of the antigenically distinct altered particles. Furthermore, the structure of the virion provides the basis for the rational development of capsid-binding inhibitors that block the genome release of CV-A6.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 カプシド) x 4
カプシド) x 4  キーワード
キーワード 機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 チェコ, 2件
チェコ, 2件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード



 PDBj
PDBj
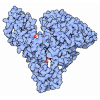




 集合体
集合体