odontoblast differentiation / Post-chaperonin tubulin folding pathway / Carboxyterminal post-translational modifications of tubulin / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Cilium Assembly / Sealing of the nuclear envelope (NE) by ESCRT-III / Intraflagellar transport / cytoskeleton-dependent intracellular transport / Formation of tubulin folding intermediates by CCT/TriC / Gap junction assembly ...odontoblast differentiation / Post-chaperonin tubulin folding pathway / Carboxyterminal post-translational modifications of tubulin / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Cilium Assembly / Sealing of the nuclear envelope (NE) by ESCRT-III / Intraflagellar transport / cytoskeleton-dependent intracellular transport / Formation of tubulin folding intermediates by CCT/TriC / Gap junction assembly / COPI-independent Golgi-to-ER retrograde traffic / natural killer cell mediated cytotoxicity / Assembly and cell surface presentation of NMDA receptors / Kinesins / GTPase activating protein binding / COPI-dependent Golgi-to-ER retrograde traffic / 細胞結合 / regulation of synapse organization / nuclear envelope lumen / cytoplasmic microtubule / Recycling pathway of L1 / MHC class I protein binding / RHOH GTPase cycle / RHO GTPases activate IQGAPs / spindle assembly / microtubule-based process / cellular response to interleukin-4 / Hedgehog 'off' state / COPI-mediated anterograde transport / Activation of AMPK downstream of NMDARs / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Recruitment of mitotic centrosome proteins and complexes / Resolution of Sister Chromatid Cohesion / Recruitment of NuMA to mitotic centrosomes / Anchoring of the basal body to the plasma membrane / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / MHC class II antigen presentation / AURKA Activation by TPX2 / Translocation of SLC2A4 (GLUT4) to the plasma membrane / RHO GTPases Activate Formins / 加水分解酵素; 酸無水物に作用; GTPに作用・細胞または細胞小器官の運動に関与 / PKR-mediated signaling / structural constituent of cytoskeleton / 紡錘体 / microtubule cytoskeleton organization / Aggrephagy / cytoplasmic ribonucleoprotein granule / HCMV Early Events / Separation of Sister Chromatids / The role of GTSE1 in G2/M progression after G2 checkpoint / microtubule cytoskeleton / Regulation of PLK1 Activity at G2/M Transition / azurophil granule lumen / double-stranded RNA binding / mitotic cell cycle / cell body / microtubule binding / 微小管 / Potential therapeutics for SARS / 細胞骨格 / calmodulin binding / 脂質ラフト / protein domain specific binding / 細胞分裂 / GTPase activity / ubiquitin protein ligase binding / Neutrophil degranulation / protein-containing complex binding / GTP binding / structural molecule activity / protein-containing complex / extracellular exosome / extracellular region / metal ion binding / 細胞核 / 細胞質基質 / 細胞質 類似検索 - 分子機能
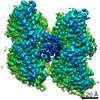
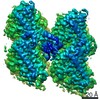
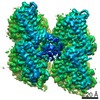
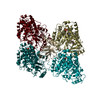
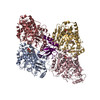
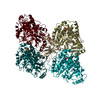
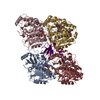
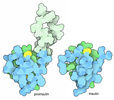
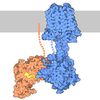
ジャーナル: Nat Commun / 年: 2019 タイトル: Structural determinants of microtubule minus end preference in CAMSAP CKK domains. 著者: Joseph Atherton / Yanzhang Luo / Shengqi Xiang / Chao Yang / Ankit Rai / Kai Jiang / Marcel Stangier / Annapurna Vemu / Alexander D Cook / Su Wang / Antonina Roll-Mecak / Michel O Steinmetz / ...著者: Joseph Atherton / Yanzhang Luo / Shengqi Xiang / Chao Yang / Ankit Rai / Kai Jiang / Marcel Stangier / Annapurna Vemu / Alexander D Cook / Su Wang / Antonina Roll-Mecak / Michel O Steinmetz / Anna Akhmanova / Marc Baldus / Carolyn A Moores / 要旨: CAMSAP/Patronins regulate microtubule minus-end dynamics. Their end specificity is mediated by their CKK domains, which we proposed recognise specific tubulin conformations found at minus ends. To ...CAMSAP/Patronins regulate microtubule minus-end dynamics. Their end specificity is mediated by their CKK domains, which we proposed recognise specific tubulin conformations found at minus ends. To critically test this idea, we compared the human CAMSAP1 CKK domain (HsCKK) with a CKK domain from Naegleria gruberi (NgCKK), which lacks minus-end specificity. Here we report near-atomic cryo-electron microscopy structures of HsCKK- and NgCKK-microtubule complexes, which show that these CKK domains share the same protein fold, bind at the intradimer interprotofilament tubulin junction, but exhibit different footprints on microtubules. NMR experiments show that both HsCKK and NgCKK are remarkably rigid. However, whereas NgCKK binding does not alter the microtubule architecture, HsCKK remodels its microtubule interaction site and changes the underlying polymer structure because the tubulin lattice conformation is not optimal for its binding. Thus, in contrast to many MAPs, the HsCKK domain can differentiate subtly specific tubulin conformations to enable microtubule minus-end recognition.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード STRUCTURAL PROTEIN (タンパク質) / Microtubule CAMSAP Calmodulin-regulated spectrum-associated proteins CKK Cryo-EM Cryo-Electron Microscopy
STRUCTURAL PROTEIN (タンパク質) / Microtubule CAMSAP Calmodulin-regulated spectrum-associated proteins CKK Cryo-EM Cryo-Electron Microscopy 機能・相同性情報
機能・相同性情報

 データ登録者
データ登録者 英国,
英国,  オランダ,
オランダ,  スイス, 6件
スイス, 6件  引用
引用

 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード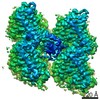

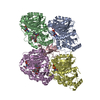
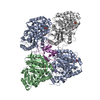

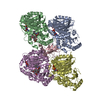
 PDBj
PDBj







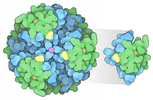















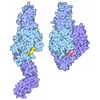


 集合体
集合体

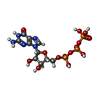

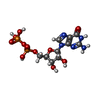
 分子量: 523.180 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H16N5O14P3 / コメント: GTP, エネルギー貯蔵分子*YM
分子量: 523.180 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H16N5O14P3 / コメント: GTP, エネルギー貯蔵分子*YM 分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg
分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM
タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM 分子量: 853.906 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C47H51NO14 / コメント: 薬剤, 化学療法薬*YM
分子量: 853.906 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C47H51NO14 / コメント: 薬剤, 化学療法薬*YM 試料調製
試料調製 電子顕微鏡撮影
電子顕微鏡撮影
 解析
解析