 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6l49 | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | H3-CA-H3 tri-nucleosome with the 22 base-pair linker DNA | |||||||||||||||||||||
 要素 要素 |
| |||||||||||||||||||||
 キーワード キーワード |  NUCLEAR PROTEIN / Chromatin (クロマチン) / Nucleosome (ヌクレオソーム) / Centromere (セントロメア) NUCLEAR PROTEIN / Chromatin (クロマチン) / Nucleosome (ヌクレオソーム) / Centromere (セントロメア) | |||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報CENP-A containing chromatin assembly / protein localization to chromosome, centromeric region / kinetochore assembly / condensed chromosome, centromeric region / establishment of mitotic spindle orientation / chromosome, centromeric region / mitotic cytokinesis / heterochromatin organization / negative regulation of tumor necrosis factor-mediated signaling pathway / negative regulation of megakaryocyte differentiation ...CENP-A containing chromatin assembly / protein localization to chromosome, centromeric region / kinetochore assembly / condensed chromosome, centromeric region / establishment of mitotic spindle orientation / chromosome, centromeric region / mitotic cytokinesis / heterochromatin organization / negative regulation of tumor necrosis factor-mediated signaling pathway / negative regulation of megakaryocyte differentiation / nucleosomal DNA binding / protein localization to CENP-A containing chromatin / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / pericentric heterochromatin / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / epigenetic regulation of gene expression / Packaging Of Telomere Ends / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / Resolution of Sister Chromatid Cohesion / Inhibition of DNA recombination at telomere / Meiotic synapsis / telomere organization / RNA Polymerase I Promoter Opening / Interleukin-7 signaling / Assembly of the ORC complex at the origin of replication / SUMOylation of chromatin organization proteins / DNAメチル化 / Condensation of Prophase Chromosomes / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / SIRT1 negatively regulates rRNA expression / Chromatin modifications during the maternal to zygotic transition (MZT) / HCMV Late Events / innate immune response in mucosa / PRC2 methylates histones and DNA / Defective pyroptosis / RHO GTPases Activate Formins / HDACs deacetylate histones / RNA Polymerase I Promoter Escape / lipopolysaccharide binding / Nonhomologous End-Joining (NHEJ) / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / NoRC negatively regulates rRNA expression / G2/M DNA damage checkpoint / B-WICH complex positively regulates rRNA expression / HDMs demethylate histones / DNA Damage/Telomere Stress Induced Senescence / Metalloprotease DUBs / PKMTs methylate histone lysines / RMTs methylate histone arginines / 遺伝的組換え / Pre-NOTCH Transcription and Translation / Activation of anterior HOX genes in hindbrain development during early embryogenesis / HCMV Early Events / Transcriptional regulation of granulopoiesis / structural constituent of chromatin / Separation of Sister Chromatids / UCH proteinases / ヌクレオソーム / antimicrobial humoral immune response mediated by antimicrobial peptide / nucleosome assembly / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / RUNX1 regulates transcription of genes involved in differentiation of HSCs / chromatin organization / Factors involved in megakaryocyte development and platelet production / HATs acetylate histones / 遺伝子発現 / Processing of DNA double-strand break ends / Senescence-Associated Secretory Phenotype (SASP) / antibacterial humoral response / Oxidative Stress Induced Senescence / killing of cells of another organism / Estrogen-dependent gene expression / defense response to Gram-negative bacterium / chromosome, telomeric region / Ub-specific processing proteases / defense response to Gram-positive bacterium / cadherin binding / protein heterodimerization activity / Amyloid fiber formation / negative regulation of cell population proliferation / chromatin binding / protein-containing complex / DNA binding / extracellular space / RNA binding / extracellular exosome / extracellular region / 核質類似検索 - 分子機能 | |||||||||||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト)synthetic construct (人工物) | |||||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 18.9 Å | |||||||||||||||||||||
 データ登録者 データ登録者 | Takizawa, Y. / Ho, C.-H. / Tachiwana, H. / Matsunami, H. / Ohi, M. / Wolf, M. / Kurumizaka, H. | |||||||||||||||||||||
| 資金援助 |  日本, 6件 日本, 6件
| |||||||||||||||||||||
 引用 引用 | ジャーナル: Structure / 年: 2020 タイトル: Cryo-EM Structures of Centromeric Tri-nucleosomes Containing a Central CENP-A Nucleosome. 著者: Yoshimasa Takizawa / Cheng-Han Ho / Hiroaki Tachiwana / Hideyuki Matsunami / Wataru Kobayashi / Midori Suzuki / Yasuhiro Arimura / Tetsuya Hori / Tatsuo Fukagawa / Melanie D Ohi / Matthias ...著者: Yoshimasa Takizawa / Cheng-Han Ho / Hiroaki Tachiwana / Hideyuki Matsunami / Wataru Kobayashi / Midori Suzuki / Yasuhiro Arimura / Tetsuya Hori / Tatsuo Fukagawa / Melanie D Ohi / Matthias Wolf / Hitoshi Kurumizaka /  要旨: The histone H3 variant CENP-A is a crucial epigenetic marker for centromere specification. CENP-A forms a characteristic nucleosome and dictates the higher-order configuration of centromeric ...The histone H3 variant CENP-A is a crucial epigenetic marker for centromere specification. CENP-A forms a characteristic nucleosome and dictates the higher-order configuration of centromeric chromatin. However, little is known about how the CENP-A nucleosome affects the architecture of centromeric chromatin. In this study, we reconstituted tri-nucleosomes mimicking a centromeric nucleosome arrangement containing the CENP-A nucleosome, and determined their 3D structures by cryoelectron microscopy. The H3-CENP-A-H3 tri-nucleosomes adopt an untwisted architecture, with an outward-facing linker DNA path between nucleosomes. This is distinct from the H3-H3-H3 tri-nucleosome architecture, with an inward-facing DNA path. Intriguingly, the untwisted architecture may allow the CENP-A nucleosome to be exposed to the solvent in the condensed chromatin model. These results provide a structural basis for understanding the 3D configuration of CENP-A-containing chromatin, and may explain how centromeric proteins can specifically target the CENP-A nucleosomes buried in robust amounts of H3 nucleosomes in centromeres. | |||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6l49.cif.gz | 834 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6l49.ent.gz | 627.9 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6l49.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/l4/6l49ftp://data.pdbj.org/pub/pdb/validation_reports/l4/6l49 | HTTPS FTP |
|---|
-関連構造データ
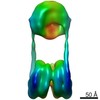

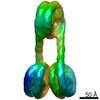
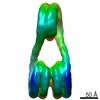
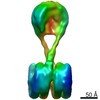





-リンク
 PDBj
PDBj













































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 5種, 24分子 AEBFLPTXCGMQUYDHNRVZKOSW
| #1: タンパク質 | 分子量: 16305.906 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CENPA / 発現宿主:  Escherichia coli (大腸菌) / 参照: UniProt: P49450 Escherichia coli (大腸菌) / 参照: UniProt: P49450#2: タンパク質 | ヒストンH4分子量: 11676.703 Da / 分子数: 6 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, ...遺伝子: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, H4FE, HIST1H4K, H4/D, H4FD, HIST1H4L, H4/K, H4FK, HIST2H4A, H4/N, H4F2, H4FN, HIST2H4, HIST2H4B, H4/O, H4FO, HIST4H4 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P62805#3: タンパク質 | 分子量: 14447.825 Da / 分子数: 6 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: HIST1H2AB, H2AFM, HIST1H2AE, H2AFA / 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P04908#4: タンパク質 | 分子量: 14217.516 Da / 分子数: 6 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: HIST1H2BJ, H2BFR / 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P06899#7: タンパク質 | ヒストンH3 / Histone H3/a / Histone H3/b / Histone H3/c / Histone H3/d / Histone H3/f / Histone H3/h / Histone ...Histone H3/a / Histone H3/b / Histone H3/c / Histone H3/d / Histone H3/f / Histone H3/h / Histone H3/i / Histone H3/j / Histone H3/k / Histone H3/l分子量: 15719.445 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H3A, H3FA, HIST1H3B, H3FL, HIST1H3C, H3FC, HIST1H3D, H3FB, HIST1H3E, H3FD, HIST1H3F, H3FI, HIST1H3G, H3FH, HIST1H3H, H3FK, HIST1H3I, H3FF, HIST1H3J, H3FJ 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: P68431 |
|---|
-DNA鎖 , 2種, 2分子 IJ
| #5: DNA鎖 | 分子量: 149214.703 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|---|
| #6: DNA鎖 | 分子量: 150401.328 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: H3-CA-H3 tri-nucleosome with the 22 base-pair linker DNA タイプ: COMPLEX / Entity ID: #1-#8 / 由来: RECOMBINANT |
|---|---|
| 分子量 | 値: 0.6 kDa/nm / 実験値: YES |
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 由来(組換発現) | 生物種: Escherichia coli (大腸菌) |
| 緩衝液 | pH: 7.8 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Talos Arctica / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TALOS ARCTICA |
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy |
| 撮影 | 電子線照射量: 25 e/Å2 フィルム・検出器のモデル: FEI FALCON II (4k x 4k) |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: (1.14_3260: phenix.real_space_refine) / 分類: 精密化 | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 対称性 | 点対称性: C2 (2回回転対称) | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 18.9 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 4999 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT |
