 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-22394 | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Functional Pathways of Biomolecules Retrieved from Single-particle Snapshots - Frame 35 - State 4 (S4) | |||||||||||||||||||||||||||
 マップデータ マップデータ | RyR-Cs2 frame 35 - state 4 (S4) | |||||||||||||||||||||||||||
 試料 試料 |
| |||||||||||||||||||||||||||
 キーワード キーワード |  ion channel (イオンチャネル) / Ca2+ channel / excitation/contraction coupling / MEMBRANE PROTEIN (膜タンパク質) ion channel (イオンチャネル) / Ca2+ channel / excitation/contraction coupling / MEMBRANE PROTEIN (膜タンパク質) | |||||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報positive regulation of sequestering of calcium ion / cyclic nucleotide binding / negative regulation of release of sequestered calcium ion into cytosol / negative regulation of insulin secretion involved in cellular response to glucose stimulus / neuronal action potential propagation / insulin secretion involved in cellular response to glucose stimulus / cell communication by electrical coupling involved in cardiac conduction / response to redox state / protein maturation by protein folding / 'de novo' protein folding ...positive regulation of sequestering of calcium ion / cyclic nucleotide binding / negative regulation of release of sequestered calcium ion into cytosol / negative regulation of insulin secretion involved in cellular response to glucose stimulus / neuronal action potential propagation / insulin secretion involved in cellular response to glucose stimulus / cell communication by electrical coupling involved in cardiac conduction / response to redox state / protein maturation by protein folding / 'de novo' protein folding / negative regulation of heart rate / negative regulation of phosphoprotein phosphatase activity / FK506 binding / positive regulation of axon regeneration / : / smooth muscle contraction / negative regulation of ryanodine-sensitive calcium-release channel activity / response to vitamin E / calcium channel inhibitor activity / regulation of cardiac muscle contraction by regulation of the release of sequestered calcium ion / protein peptidyl-prolyl isomerization / T cell proliferation / regulation of release of sequestered calcium ion into cytosol by sarcoplasmic reticulum / release of sequestered calcium ion into cytosol / Ion homeostasis / regulation of ryanodine-sensitive calcium-release channel activity / sarcoplasmic reticulum membrane / calcium channel complex / regulation of cytosolic calcium ion concentration / プロリルイソメラーゼ / peptidyl-prolyl cis-trans isomerase activity / response to hydrogen peroxide / Stimuli-sensing channels / Z disc / positive regulation of cytosolic calcium ion concentration / protein refolding / transmembrane transporter binding / signaling receptor binding / 生体膜 / 細胞質基質 / 細胞質類似検索 - 分子機能 | |||||||||||||||||||||||||||
| 生物種 |  Oryctolagus cuniculus (ウサギ) / Oryctolagus cuniculus (ウサギ) /  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||||||||||||||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.5 Å | |||||||||||||||||||||||||||
 データ登録者 データ登録者 | Dashti A / des Georges A | |||||||||||||||||||||||||||
| 資金援助 |  米国, 8件 米国, 8件
| |||||||||||||||||||||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2020 タイトル: Retrieving functional pathways of biomolecules from single-particle snapshots. 著者: Ali Dashti / Ghoncheh Mashayekhi / Mrinal Shekhar / Danya Ben Hail / Salah Salah / Peter Schwander / Amedee des Georges / Abhishek Singharoy / Joachim Frank / Abbas Ourmazd / 要旨: A primary reason for the intense interest in structural biology is the fact that knowledge of structure can elucidate macromolecular functions in living organisms. Sustained effort has resulted in an ...A primary reason for the intense interest in structural biology is the fact that knowledge of structure can elucidate macromolecular functions in living organisms. Sustained effort has resulted in an impressive arsenal of tools for determining the static structures. But under physiological conditions, macromolecules undergo continuous conformational changes, a subset of which are functionally important. Techniques for capturing the continuous conformational changes underlying function are essential for further progress. Here, we present chemically-detailed conformational movies of biological function, extracted data-analytically from experimental single-particle cryo-electron microscopy (cryo-EM) snapshots of ryanodine receptor type 1 (RyR1), a calcium-activated calcium channel engaged in the binding of ligands. The functional motions differ substantially from those inferred from static structures in the nature of conformationally active structural domains, the sequence and extent of conformational motions, and the way allosteric signals are transduced within and between domains. Our approach highlights the importance of combining experiment, advanced data analysis, and molecular simulations. #1: ジャーナル: Nat Commun / 年: 2020タイトル: Functional Pathways of Biomolecules Retrieved from Single-particle Snapshots 著者: Dashti A / des Georges A / Singharoy A / Frank J / Ourmazd A | |||||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_22394.map.gz | 230.1 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-22394-v30.xmlemd-22394.xml | 25.5 KB 25.5 KB | 表示 表示 | EMDBヘッダ |
| 画像 | 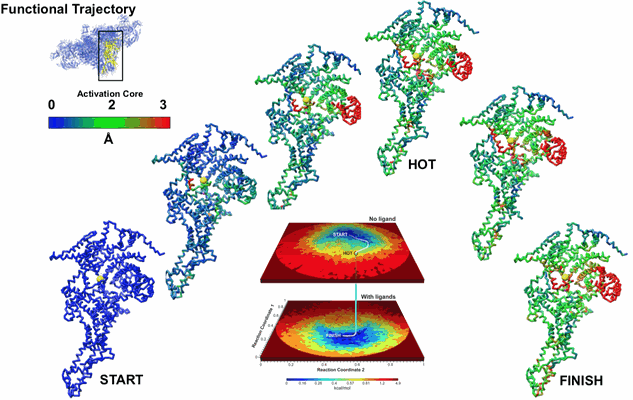 emd_22394.png emd_22394.png | 59.7 KB | ||
| Filedesc metadata | emd-22394.cif.gz | 9 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-22394ftp://ftp.pdbj.org/pub/emdb/structures/EMD-22394 http://ftp.pdbj.org/pub/emdb/structures/EMD-22394ftp://ftp.pdbj.org/pub/emdb/structures/EMD-22394 | HTTPS FTP |
-関連構造データ
















-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |





-マップ
| ファイル | ダウンロード / ファイル: emd_22394.map.gz / 形式: CCP4 / 大きさ: 244.1 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | RyR-Cs2 frame 35 - state 4 (S4) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.255 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
-添付データ
- 試料の構成要素
試料の構成要素
-全体 : Ryanodine receptor 1 bound to FKBP1B
| 全体 | 名称: Ryanodine receptor 1 bound to FKBP1B |
|---|---|
| 要素 |
|
-超分子 #1: Ryanodine receptor 1 bound to FKBP1B
| 超分子 | 名称: Ryanodine receptor 1 bound to FKBP1B / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1-#2 |
|---|---|
| 由来(天然) | 生物種: Oryctolagus cuniculus (ウサギ) |
| 分子量 | 理論値: 2.3 MDa |
-分子 #1: Peptidyl-prolyl cis-trans isomerase FKBP1B
| 分子 | 名称: Peptidyl-prolyl cis-trans isomerase FKBP1B / タイプ: protein_or_peptide / ID: 1 / コピー数: 4 / 光学異性体: LEVO / EC番号: プロリルイソメラーゼ |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 11.667305 KDa |
| 組換発現 | 生物種:  Escherichia coli (大腸菌) Escherichia coli (大腸菌) |
| 配列 | 文字列: GVEIETISPG DGRTFPKKGQ TCVVHYTGML QNGKKFDSSR DRNKPFKFRI GKQEVIKGFE EGAAQMSLGQ RAKLTCTPDV AYGATGHPG VIPPNATLIF DVELLNLE UniProtKB: Peptidyl-prolyl cis-trans isomerase FKBP1B |
-分子 #2: ryanodine receptor type 1
| 分子 | 名称: ryanodine receptor type 1 / タイプ: protein_or_peptide / ID: 2 / コピー数: 4 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Oryctolagus cuniculus (ウサギ) / 組織: thigh |
| 分子量 | 理論値: 502.246719 KDa |
| 配列 | 文字列: MGDGGEGEDE VQFLRTDDEV VLQCSATVLK EQLKLCLAAE GFGNRLCFLE PTSNAQNVPP DLAICCFTLE QSLSVRALQE MLANTVEAG VESSQGGGHR TLLYGHAILL RHAHSRMYLS CLTTSRSMTD KLAFDVGLQE DATGEACWWT MHPASKQRSE G EKVRVGDD ...文字列: MGDGGEGEDE VQFLRTDDEV VLQCSATVLK EQLKLCLAAE GFGNRLCFLE PTSNAQNVPP DLAICCFTLE QSLSVRALQE MLANTVEAG VESSQGGGHR TLLYGHAILL RHAHSRMYLS CLTTSRSMTD KLAFDVGLQE DATGEACWWT MHPASKQRSE G EKVRVGDD LILVSVSSER YLHLSTASGE LQVDASFMQT LWNMNPICSC CEEGYVTGGH VLRLFHGHMD ECLTISAADS DD QRRLVYY EGGAVCTHAR SLWRLEPLRI SWSGSHLRWG QPLRIRHVTT GRYLALTEDQ GLVVVDACKA HTKATSFCFR VSK EKLDTA PKRDVEGMGP PEIKYGESLC FVQHVASGLW LTYAAPDPKA LRLGVLKKKA ILHQEGHMDD ALFLTRCQQE ESQA ARMIH STAGLYNQFI KGLDSFSGKP RGSGPPAGPA LPIEAVILSL QDLIGYFEPP SEELQHEEKQ SKLRSLRNRQ SLFQE EGML SLVLNCIDRL NVYTTAAHFA EYAGEEAAES WKEIVNLLYE LLASLIRGNR ANCALFSTNL DWVVSKLDRL EASSGI LEV LYCVLIESPE VLNIIQENHI KSIISLLDKH GRNHKVLDVL CSLCVCNGVA VRSNQDLITE NLLPGRELLL QTNLINY VT SIRPNIFVGR AEGSTQYGKW YFEVMVDEVV PFLTAQATHL RVGWALTEGY SPYPGGGEGW GGNGVGDDLY SYGFDGLH L WTGHVARPVT SPGQHLLAPE DVVSCCLDLS VPSISFRING CPVQGVFEAF NLDGLFFPVV SFSAGVKVRF LLGGRHGEF KFLPPPGYAP CHEAVLPRER LRLEPIKEYR REGPRGPHLV GPSRCLSHTD FVPCPVDTVQ IVLPPHLERI REKLAENIHE LWALTRIEQ GWTYGPVRDD NKRLHPCLVN FHSLPEPERN YNLQMSGETL KTLLALGCHV GMADEKAEDN LKKTKLPKTY M MSNGYKPA PLDLSHVRLT PAQTTLVDRL AENGHNVWAR DRVAQGWSYS AVQDIPARRN PRLVPYRLLD EATKRSNRDS LC QAVRTLL GYGYNIEPPD QEPSQVENQS RWDRVRIFRA EKSYTVQSGR WYFEFEAVTT GEMRVGWARP ELRPDVELGA DEL AYVFNG HRGQRWHLGS EPFGRPWQSG DVVGCMIDLT ENTIIFTLNG EVLMSDSGSE TAFREIEIGD GFLPVCSLGP GQVG HLNLG QDVSSLRFFA ICGLQEGFEP FAINMQRPVT TWFSKSLPQF EPVPPEHPHY EVARMDGTVD TPPCLRLAHR (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)MPL SAAMFLSERK NPAPQCP PR LEVQMLMPVS WSRMPNHFLQ VETRRAGERL GWAVQCQDPL TMMALHIPEE NRCMDILELS ERLDLQRFHS HTLRLYRA V CALGNNRVAH ALCSHVDQAQ LLHALEDAHL PGPLRAGYYD LLISIHLESA CRSRRSMLSE YIVPLTPETR AITLFPPGR KGGNARRHGL PGVGVTTSLR PPHHFSPPCF VAALPAAGVA EAPARLSPAI PLEALRDKAL RMLGEAVRDG GQHARDPVGG SVEFQFVPV LKLVSTLLVM GIFGDEDVKQ ILKMIEPEVF TEEEEEEEEE EEEEEEEEED EEEKEEDEEE EEKEDAEKEE E EAPEGEKE DLEEGLLQMK LPESVKLQMC NLLEYFCDQE LQHRVESLAA FAERYVDKLQ ANQRSRYALL MRAFTMSAAE TA RRTREFR SPPQEQINML LHFKDEADEE DCPLPEDIRQ DLQDFHQDLL AHCGIQLEGE EEEPEEETSL SSRLRSLLET VRL VKKKEE KPEEELPAEE KKPQSLQELV SHMVVRWAQE DYVQSPELVR AMFSLLHRQY DGLGELLRAL PRAYTISPSS VEDT MSLLE CLGQIRSLLI VQMGPQEENL MIQSIGNIMN NKVFYQHPNL MRALGMHETV MEVMVNVLGG GETKEIRFPK MVTSC CRFL CYFCRISRQN QRSMFDHLSY LLENSGIGLG MQGSTPLDVA AASVIDNNEL ALALQEQDLE KVVSYLAGCG LQSCPM LLA KGYPDIGWNP CGGERYLDFL RFAVFVNGES VEENANVVVR LLIRKPECFG PALRGEGGSG LLAAIEEAIR ISEDPAR DG PGVRRDRRRE HFGEEPPEEN RVHLGHAIMS FYAALIDLLG RCAPEMHLIQ AGKGEALRIR AILRSLVPLD DLVGIISL P LQIPTL(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)NFDP RPVETLNVII PEKLDSFINK F AEYTHEKW AFDKIQNNWS YGENVDEELK THPMLRPYKT FSEKDKEIYR WPIKESLKAM IAWEWTIEKA REGEEERTEK KK TRKISQT AQTYDPREGY NPQPPDLSGV TLSRELQAMA EQLAENYHNT WGRKKKQELE AKGGGTHPLL VPYDTLTAKE KAR DREKAQ ELLKFLQMNG YAVTR(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)TPLYNL PTHRACNMFL ESYKAAWILT EDHSFEDRMI DDLSKAGEQE EEEEEVEEKK PDPLHQLVL HFSRTALTEK SKLDEDYLYM AYADIMAKSC HLEEGGENGE AEEEEVEVSF EEKEMEKQRL LYQQSRLHTR GAAEMVLQMI SACKGETGA MVSSTLKLGI SILNGGNAEV QQKMLDYLKD KKEVGFFQSI QALMQTCSVL DLNAFERQNK AEGLGMVNED G TVINRQNG EKVMADDEFT QDLFRFLQLL CEGHNNDFQN YLRTQTGNTT TINIIICTVD YLLRLQESIS DFYWYYSGKD VI EEQGKRN FSKAMSVAKQ VFNSLTEYIQ GPCTGNQQSL AHSRLWDAVV GFLHVFAHMM MKLAQDSSQI ELLKELLDLQ KDM VVMLLS LLEGNVVNGM IARQMVDMLV ESSSNVEMIL KFFDMFLKLK DIVGSEAFQD YVTDPRGLIS KKDFQKAMDS QKQF TGPEI QFLLSCSEAD ENEMINFEEF ANRFQEPARD IGFNVAVLLT NLSEHVPHDP RLRNFLELAE SILEYFRPYL GRIEI MGAS RRIERIYFEI SETNRAQWEM PQVKESKRQF IFDVVNEGGE AEKMELFVSF CEDTIFEMQI AAQISEPEGE PEADED EGM GEAAAEGAEE GAAGAEGAAG TVAAGATARL AAAAARALRG LSYRSLRRRV RRLRRLTARE AATALAALLW AVVARAG AA GAGAAAGALR LLWGSLFGGG LVEGAKKVTV TELLAGMPDP TSDEVHGEQP AGPGGDADGA GEGEGEGDAA EGDGDEEV A GHEAGPGGAE GVVAVADGGP FRPEGAGGLG DMGDTTPAEP PTPEGSPILK RKLGVDGEEE ELVPEPEPEP EPEPEKADE ENGEKEEVPE APPEPPKKAP PSPPAKKEEA GGAGMEFWGE LEVQRVKFLN YLSRNFYTLR FLALFLAFAI NFILLFYKVS DSPPGEDDM EGSAAGDLAG AGSGGGSGWG SGAGEEAEGD EDENMVYYFL EESTGYMEPA LWCLSLLHTL VAFLCIIGYN C LKVPLVIF KREKELARKL EFDGLYITEQ PGDDDVKGQW DRLVLNTPSF PSNYWDKFVK RKVLDKHGDI FGRERIAELL GM DLASLEI TAHNERKPDP PPGLLTWLMS IDVKYQIWKF GVIFTDNSFL YLGWYMVMSL LGHYNNFFFA AHLLDIAMGV KTL RTILSS VTHNGKQLVM TVGLLAVVVY LYTVVAFNFF RKFYNKSEDE DEPDMKCDDM MTCYLFHMYV GVRAGGGIGD EIED PAGDE YELYRVVFDI TFFFFVIVIL LAIIQGLIID AFGELRDQQE QVKEDMETKC FICGIGSDYF DTTPHGFETH TLEEH NLAN YMFFLMYLIN KDETEHTGQE SYVWKMYQER CWDFFPAGDC FRKQYEDQLS |
-分子 #3: ZINC ION
| 分子 | 名称: ZINC ION / タイプ: ligand / ID: 3 / コピー数: 4 / 式: ZN |
|---|---|
| 分子量 | 理論値: 65.409 Da |
-分子 #4: CALCIUM ION
| 分子 | 名称: CALCIUM ION / タイプ: ligand / ID: 4 / コピー数: 4 / 式: CA |
|---|---|
| 分子量 | 理論値: 40.078 Da |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | 3D array |
-試料調製
| 濃度 | 8 mg/mL |
|---|---|
| 緩衝液 | pH: 7.4 |
| グリッド | モデル: UltrAuFoil / 材質: GOLD / 支持フィルム - 材質: GOLD / 支持フィルム - トポロジー: HOLEY ARRAY |
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 100 % / チャンバー内温度: 277 K / 装置: FEI VITROBOT MARK IV |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI POLARA 300 |
|---|---|
| 電子線 | 加速電圧: 300 kV / 電子線源: FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELDBright-field microscopy |
| 試料ステージ | 試料ホルダーモデル: GATAN ULTST ULTRA LOW TEMPERATURE SINGLE TILT HELIUM COOLING HOLDER ホルダー冷却材: NITROGEN |
| 撮影 | フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 検出モード: COUNTING / 平均電子線量: 50.0 e/Å2 |
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: EMDB MAP EMDB ID: 詳細: RyR1-Cs2 (EGTA-only) |
|---|---|
| 初期 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
| 最終 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
| 最終 再構成 | 想定した対称性 - 点群: C4 (4回回転対称) / 解像度のタイプ: BY AUTHOR / 解像度: 4.5 Å / 解像度の算出法: OTHER 詳細: RESMAP and visual inspection. FSC not possible as no half-sets are available with the manifold embedding method 使用した粒子像数: 791956 |

-原子モデル構築 1
| 初期モデル | PDB ID: Chain - Source name: PDB / Chain - Initial model type: experimental model |
|---|---|
| 詳細 | fitted to a model domain by domain with the rigid-body fit function in COOT 71, using multiple starting models to avoid model bias (PDB ID: 5TB4, 5T9R, 5TAP, 5T9V, 5TAL, 5TAQ) 22. The models were then refined in real-space using phenix.real_space_refine |
| 精密化 | 空間: REAL / プロトコル: OTHER |
| 得られたモデル |  PDB-7jmh: |

