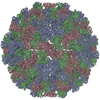
Journal: J Mol Biol / Year: 1986 Title: Structure and assembly of turnip crinkle virus. I. X-ray crystallographic structure analysis at 3.2 A resolution. Abstract: The structure of turnip crinkle virus has been determined at 3.2 A resolution, using the electron density of tomato bushy stunt virus as a starting point for phase refinement by non-crystallographic ...The structure of turnip crinkle virus has been determined at 3.2 A resolution, using the electron density of tomato bushy stunt virus as a starting point for phase refinement by non-crystallographic symmetry. The structures are very closely related, especially in the subunit arm and S domain, where only small insertions and deletions and small co-ordinate shifts relate one chain to another. The P domains, although quite similar in fold, are oriented somewhat differently with respect to the S domains. Understanding of the structure of turnip crinkle virus has been important for analyzing its assembly, as described in an accompanying paper.
History
Deposition
Aug 8, 2011
Deposition site: PDBE / Processing site: PDBE
Revision 1.0
Feb 8, 2012
Provider: repository / Type: Initial release
Revision 1.1
Mar 4, 2015
Group: Database references
Revision 1.2
Feb 3, 2016
Group: Derived calculations
Revision 1.3
Jun 28, 2017
Group: Data collection / Category: diffrn_source Item: _diffrn_source.pdbx_wavelength_list / _diffrn_source.type
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
Basic information Components
Components Capsid
Capsid  Keywords
Keywords Function and homology information
Function and homology information
 Authors
Authors Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads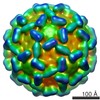




 PDBj
PDBj

 Assembly
Assembly



